Volume 10, Number 3—March 2004
Research
Amoebae-resisting Bacteria Isolated from Human Nasal Swabs by Amoebal Coculture
Abstract
Amoebae feed on bacteria, and few bacteria can resist their microbicidal ability. Amoebal coculture could therefore be used to selectively grow these amoebae-resisting bacteria (ARB), which may be human pathogens. To isolate new ARB, we performed amoebal coculture from 444 nasal samples. We recovered 7 (1.6%) ARB from 444 nasal swabs, including 4 new species provisionally named Candidatus Roseomonas massiliae, C. Rhizobium massiliae, C. Chryseobacterium massiliae, and C. Amoebinatus massiliae. The remaining isolates were closely related to Methylobacterium extorquens, Bosea vestrii, and Achromobacter xylosoxidans. Thus, amoebal coculture allows the recovery of new bacterial species from heavily contaminated samples and might be a valuable approach for the recovery of as-yet unrecognized emerging pathogens from clinical specimens.
Free-living amoebae have been isolated worldwide from soil, water, air (1), and from the noses of healthy persons (2,3). Most amoebae feed on bacteria, and few bacteria can resist digestion by the amoebae after being internalized. Amoebal coculture could therefore be used to selectively grow these amoebae-resisting bacteria (ARB) and clean the samples of other more rapidly growing species that generally overwhelm the agar plates. ARB include established agents of pneumonia such as Legionella spp (4). and Chlamydophila pneumoniae (5), and potential human pathogens, such as Parachlamydia spp (6,7). and Legionella-like amoebal pathogens (LLAPs) (8).
Human exposure to Parachlamydia spp. has been especially well demonstrated. Indeed, two of the eight Parachlamydiaceae strains isolated to date were found within amoebae recovered from the nasal mucosa (3). However, the prevalence of this potential human pathogen, of other ARB, and of their amoebal host on the nasal mucosa remains to be determined. The fact that Parachlamydia is endosymbiotic at 30°C and lytic at 37°C suggests that the bacteria present within amoebae in the cooler upper respiratory tract may be liberated when the amoebae reach the warmer lower respiratory tract (9). This simultaneous liberation of bacteria may overwhelm the human immune response and explain the role of Parachlamydia as an agent of community-acquired pneumonia (6) and of aspiration pneumonia (10).
We hypothesized that, for any ARB, nasal colonization may be a first step towards lower respiratory tract infection. Moreover, ARB may use free-living amoebae as a training ground for the selection of virulence traits, as demonstrated by Cryptococcus neoformans (11). Thus, virulent capsular strains were phagocytosed by and replicated in Acanthamoeba castellanii, leading to amoebal death, while the nonvirulent acapsular strains were killed (11). The fact that free-living amoebae may promote the expression of virulence traits was also well demonstrated for L. pneumophila (12,13) and Mycobacterium avium (14). A growing body of evidence support the hypothesis that free-living amoebae play a role in the adaptation of L. pneumophila to life within other phagolysosomal cells, such as human macrophages (15–18). Moreover, the growth of M. avium in amoebae, which results in enhanced entry into both amoebae and macrophages (14), further supports the hypothesis that adaptation to life within human macrophages may be acquired after exposure to environmental predators such as free-living amoebae. Thus, free-living amoebae may be considered as an evolutionary crib for potential emerging human pathogens.
We intended to isolate new ARB from nasal swabs and to compare the prevalence of free-living amoebae and of ARB, such as Parachlamydia acanthamoebae, in healthy participants, homeless persons, and hospitalized patients. We tried to recover ARB and free-living amoebae by using amoebal coculture and amoebal enrichment, respectively, with any recovered amoebae subsequently examined for the presence of ARB.
Patients and Study Design
Nasal swabs were taken from 100 patients hospitalized in the infectious diseases unit of the Hôpital Nord of Marseille, France, and from 244 homeless persons. In addition, nasal swabs were taken from 100 healthy participants. All participants took part freely in the study and gave their written informed consent. Amoebal coculture was performed for all 444 samples, while amoebal enrichment on nonnutrient agar was restricted to the samples taken from the 100 healthy volunteers and the 100 hospitalized patients.
Broth and Media
Broth and media were slightly modified from the method of T.J. Rowbotham (19). Peptone yeast-extract glucose (PYG): 100 g proteose peptone (Difco, Sparks, MD), 10 g yeast-extract (Merck, Darmstadt, Germany), 4.9 g MgSO4.7H2O, 5 g sodium citrate.2H2O, 0.1 g Fe(NH4)2 (SO4)2.6H2O, 1.7 g KH2PO4, 1.97 g Na2HPO4.7H20, 45 g glucose, and 0.295 g CaCl2 in 5 L of distilled water. Page’s modified Neff’s amoebae saline (PAS): 120 mg NaCl, 4 mg MgSO4.7H2O, 4 mg CaCl2.2H2O, 142 mg Na2HPO4, and 136 mg KH2PO4 in 1 L of distilled water. Nonnutritive 1.5 g agar (Research Organics, Cleveland, OH) was diluted in 100 mL of PAS. All media were autoclaved 15 minutes at 121°C, and broths were also filtered through a 0.22-μm pore membrane (Corning, New York, NY).
Amoebal Microplates
An Acanthamoeba polyphaga strain, Linc-AP1, was grown at 28°C in 150-cm2 cell culture flasks (Corning) with 30 mL PYG. After 5 days, the amoebae were harvested and centrifuged at 179 X g (1,000 rpm) for 10 minutes. The supernatant was removed, and the amoebal pellet was resuspended in 50 mL of PAS. Centrifugation and resuspension steps were repeated twice. After the last centrifugation, the amoebae were resuspended in PAS, and 1 mL of 5x105 A. polyphaga/mL was distributed in each well of a 12-well Costar microplate (Corning).
Nasal Samples
Nasal swabs were vortexed for 30 seconds in 1 mL of PAS in individual sterile tubes. The suspension was centrifuged at 179 X g (1,000 rpm) for 10 minutes. Two hundred and fifty microliters of supernatant was spread onto amoebal microplates. The pellet was spread on nonnutritive agar plates previously flooded with a turbid suspension of viable Enterobacter aerogenes in PAS, drained, and incubated at 28°C for a few hours.
Amoebal Coculture
The microplates were centrifuged at 2,879 X g (4,000 rpm) for 30 minutes and incubated at 32°C in a humidified CO2 atmosphere. The amoebal cocultures were subcultured on fresh amoebae on days 4 and 7 (samples from controls and patients) or on day 6 (samples from homeless persons). Amoebal cocultures were examined regularly for amoebal lysis. When amoebal lysis was observed and at the time of subculture, the cultures were screened for intra-amoebal bacteria. This screening was achieved by gently shaking the microplates to suspend the amoebae. Then 150 μL of the suspension was cytocentrifuged at 150 X g (800 rpm) for 10 minutes, and slides were stained with Gimenez (20). Fuchsin-stained bacteria easily could be seen within the malachite green-stained amoebae. When Gimenez-stained bacteria were observed, the culture was subcultured on BCYE agar (BioMérieux, Marcy L’Etoile, France) and sheep blood agar (BioMérieux) and on amoebal microplates without antibiotics or with vancomycin (10 μg/mL) or colistin (500 U/mL).
Amoebal Enrichment
Nonnutritive agar plates not supplemented with amphotericin B were incubated at 28°C in a humidified CO2 atmosphere and examined daily for amoebae-like cells. When positive, these amoebae-like cells were subcultured on nonnutritive agar plates seeded with viable or UV–inactivated Enterobacter aerogenes. After several subcultures on inactivated E. aerogenes, recovered amoebae were tentatively grown in axenic PYG broth with or without antimicrobials.
Phenotypic Characterization
Microscopic properties of the isolated bacteria were studied by Gram and Gimenez staining. To confirm that the isolated strains were the Gimenez-stained bacteria, 5 X 105 A. polyphaga/mL suspensions were injected with the isolated strain and incubated at 32°C. After 4 days’ incubation, culture broth was cytocentrifuged and stained with Gimenez and Gram, as described above.
Genotypic and Phylogenetic Characterization
DNA was extracted according to the manufacturer’s instructions by using Chelex Resin (BioRad, Hercules, CA) or FastDNA kit (Bio101, Carlsbad, CA) and FastPrep120 grinder (Bio101), depending on the time of strain recovery. Polymerase chain reaction (PCR) amplification of the 16S rDNA gene was performed by using the fD1 and rP2 primers (21) and Taq DNA polymerase (GibcoBRL, Life technologies, Rockville, MD), according to manufacturer instructions. The success of the amplification was determined by electrophoresis in 1% agarose gel of PCR products stained with ethidium bromide. PCR products were purified by using the QIAquick PCR purification kit (Qiagen, Courtaboeuf, France). Sequencing was performed with the dRhodamine Terminator Cycle sequencing Ready Reaction with one of nine different primers and with AmpliTaq DNA (Perkin-Elmer Biosystems, Warrington, England), according to the manufacturer’s instructions. Sequences were determined on a 3100 ABI Prism automated sequencer (Applied Biosystems, Courtaboeuf, France), according to the manufacturer’s instructions. Sequences derived from each primer were aligned, compared, and combined in a single 16S rDNA sequence, using the Autoassembler software version 2.1 (Applied Biosystems). The validity of the sequence obtained was assessed by comparison with two additional sequences obtained similarly but from two other PCRs of the same template DNA. The sequences were compared with sequences available in the GenBank database in October 2002 by using the BLAST 2.2.2 program available on the NCBI Web site (www.ncbi.nlm.nih.gov) (22). When the closest sequence was that of an as-yet unpublished bacterial species, the 16S rDNA sequence homology of the prototype strain of the closest published bacterial species was also identified (Table). The 16S rDNA sequences of the isolates were aligned with the sequences of the best blast hits and with those of the closest relative type strains of each isolate. Sequences were edited by removal of the longer 5′ and 3′ ends so that their lengths matched that of the shortest sequence. The 16S rDNA homology of the edited sequences was then analyzed by the distance matrix program of the MEGA software (32). We considered 16S rDNA sequence similarity to be that of the closest sequence in GenBank of more than 99% and 97% for identification at the species and genus level, respectively (33). With these sequences, neighbor-joining (Kimura 2-parameter), minimum evolution (Kimura 2-parameter), and parsimony (standard parsimony) trees were constructed with the MEGA software (32).
Electron Microscopy
For some strains, reinoculation procedure was also performed onto 5x105 A. polyphaga/mL suspension in 50-cm3 flasks (Corning). Four days’ coculture was harvested, washed in monophosphate buffer (pH 7.2), and fixed in 4% glutaraldehyde. Fixed samples were washed overnight in monophosphate buffer (pH 7.2), and then fixed for 1 hour at 4°C with 1% osmium tetroxyde. Dehydration was performed by successive washes in increasing acetone concentrations (25%–100%). After incubating the preparations for 1 hour in a vol/vol suspension of acetone/epon and then overnight in epon, they were embedded in Araldite (Fluka, St Quentin Fallavier, France). Thin sections were cut from embedded blocks with an Ultracut microtome (Reichert-Leica, Marseille, France), deposited on copper grids coated with formvar (Sigma-Aldrich, Taufkirchen, Germany), and stained 10 minutes with a solution of methanol-uranyl acetate and lead nitrate with sodium citrate in water. Grids were examined with a Morgagni 268D electron microscope (Philips, Eindhoven, the Netherlands).
Statistical Analysis
The mean number of strains isolated from ill hospitalized patients was compared to the mean number of strains isolated from the other participants (healthy participants and homeless cohort) by using the student t test (STATA software version 7.0., Stata Corporation, College Station, TX).
Amoebal Enrichment
We observed trophozoites growing on nonnutrient agar injected into 1 of 200 samples but failed in propagating them due to fungal contamination, despite subcultures on nonnutrient agar not supplemented and supplemented with amphotericin B (2 μg/mL). Thus, no free-living amoebae could be successfully recovered. Fungal contamination was frequent, occurring in 61% of cultures from patients and 41% of cultures from control samples. These fungi were not identified.
Amoebal Coculture
By amoebal coculture, we recovered seven (1.6%) strains of ARB from nasal swabs of 4 of 444 participants. Five isolates were recovered from 100 patients hospitalized in the infectious diseases unit of the Hôpital Nord of Marseilles, whereas only two isolates were recovered from the 344 other participants (244 homeless and 100 healthy volunteers; p = 0.017). All ARB were gram-negative bacilli. No amoebae-resisting fungi were detected. On the basis of 16S rDNA sequence analysis, the isolated ARB were assigned to alphaproteobacteria (n = 4), betaproteobacteria (n = 1), and the Bacteroides-Cytophaga-Flexibacter group (n = 2). The Table shows the taxonomy and the 16S rDNA homology with GenBank sequences of the seven isolated ARB. When we used a 16S rDNA sequence similarity of more than 99% and 97% for identification at the species and genus level, respectively (33), four (57%) ARB belonged to new species (isolates 1, 4, 6, and 7), including one that also corresponded to a new genus (isolate 6).
Figure 1
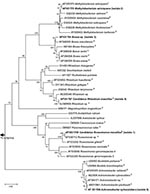
Figure 1. 16S rDNA tree showing relationship of isolates 1 to 5 with related alpha- and beta-proteobacteria. The tree was constructed by using the neighbor-joining method, based on the nearly complete sequence (1,289...
Isolate 1, Candidatus Roseomonas massiliae, was recovered from a 44-year-old healthy homeless man. The 16S rDNA sequence of isolate 1 shared on BLAST analysis 99.9% homology with a Roseomonas sp. implicated in a catheter-related bacteremia (X.Y. Han, unpub. data) and 98.8% with R. gilardii strain ATCC 49956T (Table). Genetic analysis confirmed that isolate 1 belonged to the genus Roseomonas, as it shares 93.9%–98.9% 16S rDNA sequence homology with Roseomonas spp. and 94.3%–84.6% with other Methylobacteriaceae, respectively. Isolate 1 clustered with Roseomonas spp. by phylogenetic analysis of the 16S rDNA gene (Figure 1). Thus, bootstrap values of 98%, 99%, and 90% in neighbor-joining, minimum evolution, and parsimony analyses, respectively, supported the fork separating Roseomonas spp. (including isolate 1) from their closest relatives, Craurococcus roseus and Paracraurococcus ruber. More importantly, the isolate was clearly different from the type strain neighbor (R. gilardii), with bootstrap values of 100% in neighbor-joining, minimum evolution, and parsimony trees. Isolate 1 has been deposited in the Collection de l’Institut Pasteur, Paris, France, as Candidatus Roseomonas massiliae strain CIP 107751T.
Isolate 2 was recovered from a 27-year-old homeless man with chronic cough, headache, and polylymphadenopathy. The 16S rDNA sequence of isolate 2 shared on BLAST analysis 99.3% homology with Methylobacterium extorquens strain JCM 2802T (Table). Genetic analysis confirmed that isolate 2 is an additional strain of M. extorquens as it shared 99.3% 16S rDNA sequence homology with M. extorquens strain JCM 2802T and 94.6%–99% with the other Methylobacterium spp. Moreover, isolate 2 clustered with Methylobacterium spp. by phylogenetic analysis of the 16S rDNA gene (Figure 1).
Figure 2
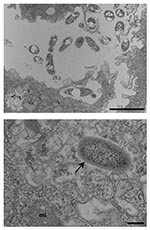
Figure 2. A. Bosea sp. (isolate 3) in the process of being phagocytized by Acanthamoeba polyphaga (arrows) and in the extracellular media, as seen on electron microscopy. 8,900 x magnification. Bar represents 200...
Isolate 3 was recovered from a 48-year-old HIV-positive man with cerebral toxoplasmosis and severe pneumonia attributed to Streptococcus pneumoniae. The 16S rDNA sequence of isolate 3 shared on BLAST analysis 99.2% homology with Bosea vestrii strain 34635T (Table). Genetic analysis confirmed that isolate 3 belonged to the genus Bosea, as it shares 98%–99% 16S rDNA sequence homology with Bosea spp. Isolate 3 clustered with Bosea spp. by phylogenetic analysis of the 16S rDNA gene (Figure 1). However, it clustered with B. massiliensis, with bootstrap values separating them (isolate 3 and B. massiliensis) from the other Bosea spp. of 100%, 56%, and 100% in neighbor-joining, minimum evolution, and parsimony analyses, respectively. The morphology of isolate 3 within A. polyphaga, as seen by electron microscopy, is shown in Figure 2.
Isolate 4, Candidatus Rhizobium massiliae, was recovered from an 80-year-old woman with pacemaker-associated endocarditis. The 16S rDNA sequence of isolate 4 shared on BLAST analysis 98.7% homology with Rhizobium larrymoorei strain 3–10 (Table). Although the isolate shared greater homology with sequences annotated as Rhizobium sp., Agrobacterium sp., or A. tumefaciens (A. tumefaciens was renamed R. radiobacter [34]), it shared 92.4% sequence homology with R. radiobacter strain IFO 13257T (accession no. D14501). Isolate 4 clustered with two as yet unclassified strains of Rhizobium sp. by phylogenetic analysis of the 16S rDNA gene with a node separating them from the closest type strain neighbor, R. larrymoorei supported by bootstrap values of 100%, 50%, and 100% in neighbor-joining, minimum evolution and parsimony trees, respectively. Isolate 4 has been deposited in the Collection de l’Institut Pasteur, Paris, France, as Candidatus Rhizobium massiliae strain CIP 107749T.
Isolate 5 (like isolates 4 and 7) was recovered from an 80-year-old woman with a pacemaker-associated endocarditis. The 16S rDNA sequence of isolate 5 shared on BLAST analysis 99.5% homology with Achromobacter xylosoxidans strain ATCC 9220T (Table). Genetic analysis confirmed that isolate 5 belonged to the genus Achromobacter, as it shares a 98.4%–99.5% 16S rDNA sequence homology with Achromobacter spp. and 97.6%–97.9% with Bordetella spp. Isolate 5 clustered with Achromobacter spp. by phylogenetic analysis of the 16S rDNA gene (Figure 1). Thus, bootstrap values of 100% in neighbor-joining, minimum evolution, and parsimony trees supported the fork’s separating the Achromobacter spp. (including isolate 5) from Bordetella spp.
Figure 3

Figure 3. 16S rDNA tree showing relationship of isolates 6 and 7 with related Flavobacteriaceae. The tree was constructed by using the neighbor-joining method, based on the nearly complete sequence (1,283 nt) of...
Isolate 6, Candidatus Amoebinatus massiliae, (like isolate 3) was recovered from a 48-year-old HIV-positive man with cerebral toxoplasmosis and severe pneumonia attributed to S. pneumoniae. The 16S rDNA sequence of isolate 6 shared on BLAST analysis 99.2% homology with that of a pathogenic strain belonging to an unpublished new genus and 95.3% with Chryseobacterium scophtalmum strain LMG 13028T (Table). On genetic analysis, isolate 6 shared 93.1%–96.0% 16S rDNA sequence homology with Chryseobacterium spp. and 81.5%–90.0% with other Flavobacteriaceae, respectively. Phylogenetic analysis of the 16S rDNA gene also suggested that isolate 6 belonged to a new genus, related to the Chryseobacterium spp. (Figure 3). Thus, bootstrap values of 100%, 100%, and 34% in neighbor-joining, minimum evolution, and parsimony analyses, respectively, supported the fork’s separating both Chryseobacterium-like bacteria (including isolate 6) from their closest relatives. Isolate 6 has been deposited in the Collection de l’Institut Pasteur, Paris, France, as Candidatus Amoebinatus massiliae strain CIP 107750T.
Isolate 7, Candidatus Chryseobacterium massiliae, (like isolates 4 and 5) was recovered from an 80-year-old woman with pacemaker-associated endocarditis. The 16S rDNA sequence of isolate 7 shared on BLAST analysis 98.0% homology with that of a strain isolated from the lower respiratory tract of a cystic fibrosis patient (35) and 96.3% with Chryseobacterium scophtalmum strain LMG 13028T (Table). On genetic analysis, isolate 7 shared 92.5%–97.1% 16S rDNA sequence homology with Chryseobacterium spp. and 81.8%–90.3% with other Flavobacteriaceae, respectively. Phylogenetic analysis of the 16S rDNA gene (Figure 3) suggested that isolate 7 represented a new species of Chryseobacterium (Figure 3). Thus, bootstrap values of 100%, 100%, and 97% in neighbor-joining, minimum evolution, and parsimony analyses, respectively, supported the fork’s separating isolate 6 from its closest neighbor, an unpublished species of Chryseobacterium. The morphologic features of isolate 7 within A. polyphaga, as seen by electron microscopy, is shown in Figure 2. Isolate 7 has been deposited in the Collection de l’Institut Pasteur, Paris, France, as Candidatus Chryseobacterium massiliae strain CIP 107752T.
Recovery of Methylobacteriaceae, Rhizobiaceae, Alcaligenaceae, and Flavobacteriacae by amoebal coculture shows that these clades should be added to the growing number of ARB. Our study also shows that humans are exposed to unknown ARB and that the amoebal coculture is an effective tool for the recovery of new species from contaminated samples.
Until this study, the ARB included Legionella spp (4,36), Chlamydiales (3,5), Bradyrhizobiaceae (37), Rickettsiales (38), Listeria monocytogenes (39), M. avium (40), Procabacter acanthamoeba, a betaproteobacteria (41), and members of the Bacteroides-Cytophaga-Flexibacter group (Flavobacterium spp. and Amoebophilus asiaticus) (42). In this study, we recovered additional alphaproteobacteria including a strain of Methylobacterium extorquans, a Bosea sp., a new Rhizobium sp., provisionally named Candidatus Rhizobium massiliae, and a new Roseomonas sp., named C. Roseomonas massiliae. We also recovered a strain of Achromobacter xylosoxidans (betaproteobacteria) and two new Flavobacteriacae related to the Chryseobacterium spp., provisionally named C. Chryseobacterium massiliae and C. Amoebinatus massiliae. Whether these species use the free-living amoebae as a reservoir and whether they play a role as human pathogens remain to be defined.
ARB in nasal passages of patients and homeless persons are important because they demonstrate the exposure of humans to these bacteria. From the nasal mucosa, ARB may spread to the lower respiratory tract, where they might cause bronchitis and pneumonia, being somewhat adapted to intracellular life. The fact that ARB were more likely to be recovered from patients hospitalized in the infectious disease department than from the other patients sampled is of interest. Further studies will be needed to confirm this statistically significant association and to determine whether patients were colonized with ARB during their hospitalization, whether colonization with ARB may be a marker of exposition to another pathogen involved in the infectious process that lead to hospitalization, or whether the isolated ARB may itself be the etiologic agent of the infectious process that led to hospitalization.
Our study also shows that amoebal coculture is a cell culture system that may be used to recover new bacterial species from heavily contaminated clinical or environmental samples. Indeed, by grazing on bacteria, the free-living amoebae will clean the samples from most rapid growing species that generally overwhelm the agar plates (for example Enterobacteriaceae, Staphylococci, Streptococci, and Neisseriaceae in a nasal sample). Thus, using that technique, Rowbotham was able to grow L. pneumophila from human feces (43). Moreover, the amoebal coculture is a cell culture system that may be performed in the absence of antimicrobials and is thus especially suitable for recovery of new bacterial species of unknown antimicrobial susceptibility. Relative to a broad amplification strategy for microbial rDNA found in nasal washings, amoebal coculture has several advantages. First, it makes a strain available; its phenotype, genotype, antibiotic susceptibility, and virulence may then be further characterized. Second, a broad amplification strategy is unlikely to be successful in obtaining a clear sequence from such heavily contaminated sample. As amoebal resistance may be associated with virulence (11–16), the amoebal coculture might have the additional advantage of selectively growing potential human pathogens. The main limitations of this technique are the decreased viability of the amoebal cells and their encystment at high incubation temperature, which do not allow the recovery of bacteria requiring a temperature >37°C. Another limitation is that, contrary to amoebal enrichment, amoebal coculture did not recover the amoebal host, if any, present in the sample. In addition, this method, like other cell cultures, is associated with a substantial laboratory workload.
We did not isolate Parachlamydia spp., which may reflect the scarcity of sources or the low or nonexistent exposure to this potential human pathogen in the area where the survey was performed. This finding may also explain the low prevalence of Parachlamydia-associated community-acquired pneumonia that we observed in a recent study (44).
In conclusion, amoebal coculture allows new bacterial species to be isolated from heavily contaminated human samples and might be a valuable approach in identifying potential, as-yet unrecognized human pathogens from nonsterile clinical samples, such as upper and lower respiratory tract specimen.
Dr. Greub is a specialist in medical microbiology and infectious disease. He currently works at the Institute of Microbiology of the University of Lausanne, Switzerland. His research focuses on Parachlamydia acanthamoebae and other emerging intracellular bacteria.
Acknowledgment
We thank the Swiss National Science Foundation for funding the postdoctoral fellowship of Gilbert Greub; K. Johnston for reviewing the manuscript; P. Brouqui for allowing us to swab the noses of 100 patients hospitalized in the Division of Infectious Diseases, Hôpital Nord, Marseille; and T.J. Rowbotham for kindly providing an Acanthamoeba polyphaga strain, Linc-AP1.
References
- Rodriguez-Zaragoza S. Ecology of free-living amoebae. Crit Rev Microbiol. 1994;20:225–41. DOIPubMedGoogle Scholar
- Michel R, Hauröder-Philippczyk B, Muller KD, Weishaar I. Acanthamoeba from human nasal mucosa infected with an obligate intracellular parasite. Eur J Protistol. 1994;30:104–10.
- Amann R, Springer N, Schönhuber W, Ludwig W, Schmid EN, Muller KD, Obligate intracellular bacterial parasites of Acanthamoebae related to Chlamydia spp. Appl Environ Microbiol. 1997;63:115–21.PubMedGoogle Scholar
- Rowbotham TJ. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J Clin Pathol. 1980;33:1179–83. DOIPubMedGoogle Scholar
- Essig A, Heinemann M, Simnacher U, Marre R. Infection of Acanthamoeba castellanii by Chlamydia pneumoniae. Appl Environ Microbiol. 1997;63:1396–9.PubMedGoogle Scholar
- Birtles RJ, Rowbotham TJ, Storey C, Marrie TJ, Raoult D. Chlamydia-like obligate parasite of free-living amoebae. Lancet. 1997;349:925–6. DOIPubMedGoogle Scholar
- Greub G, Raoult D. Parachlamydiaceae: potential emerging pathogens. Emerg Infect Dis. 2002;8:625–30.PubMedGoogle Scholar
- Marrie TJ, Raoult D, La Scola B, Birtles RJ, de Carolis E. Legionella-like and other amoebal pathogens as agents of community-acquired pneumonia. Emerg Infect Dis. 2001;7:1026–9. DOIPubMedGoogle Scholar
- Greub G, La Scola B, Raoult D. Parachlamydia acanthamoebae is endosymbiotic or lytic for Acanthamoeba polyphaga depending on the incubation temperature. Ann N Y Acad Sci. 2003;990:628–34. DOIPubMedGoogle Scholar
- Greub G, Boyadjiev I, La Scola B, Raoult D, Martin C. Serological hint suggesting that Parachlamydiaceae are agents of pneumonia in polytraumatized intensive-care patients. Ann N Y Acad Sci. 2003;990:311–9. DOIPubMedGoogle Scholar
- Steenbergen JN, Shuman HA, Casadevall A. Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc Natl Acad Sci U S A. 2001;98:15245–50. DOIPubMedGoogle Scholar
- Rowbotham TJ. Current views on the relationships between amoebae, legionellae and man. Isr J Med Sci. 1986;22:678–89.PubMedGoogle Scholar
- Cirillo JD, Falkow S, Tompkins LS. Growth of Legionella pneumophila in Acanthamoeba castellanii enhances invasion. Infect Immun. 1994;62:3254–61.PubMedGoogle Scholar
- Cirillo JD, Falkow S, Tompkins LS, Bermudez LE. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect Immun. 1997;65:3759–67.PubMedGoogle Scholar
- Cianciotto NP, Fields BS. Legionella pneumophila mip gene potentiates intracellular infection of protozoa and human macrophages. Proc Natl Acad Sci U S A. 1992;89:5188–91. DOIPubMedGoogle Scholar
- Gao LY, Harb OS, Abu Kwaik Y. Utilization of similar mechanisms by Legionella pneumophila to parasitize two evolutionarly distant host cells, mammalian macrophages and protozoa. Infect Immun. 1997;65:4738–46.PubMedGoogle Scholar
- Stone BJ, Brier A, Kwaik YA. The Legionella pneumophila prp locus; required during infection of macrophages and amoebae. Microb Pathog. 1999;27:369–76. DOIPubMedGoogle Scholar
- Segal G, Shuman HA. Legionella pneumophila utilizes the same genes to multiply within Acanthamoeba castellanii and human macrophages. Infect Immun. 1999;67:2117–24.PubMedGoogle Scholar
- Rowbotham TJ. Isolation of Legionella pneumophila from clinical specimens via amoebae, and the interaction of those and other isolates with amoebae. J Clin Pathol. 1983;36:978–86. DOIPubMedGoogle Scholar
- Gimenez DF. Staining rickettsiae in yolk-sac cultures. Stain Technol. 1964;39:135–40.PubMedGoogle Scholar
- Weisburg WG, Barns SM, Pelletier DA, Lane DJ. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol. 1991;173:697–703.PubMedGoogle Scholar
- Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402. DOIPubMedGoogle Scholar
- Rihs JD, Brenner DJ, Weaver RE, Steigerwalt AG, Hollis DG, Yu VL. Roseomonas, a new genus associated with bacteremia and other human infections. J Clin Microbiol. 1993;31:3275–83.PubMedGoogle Scholar
- Urakami T, Komagata K. Protomonas, a new genus of facultatively methylotrophic bacteria. Int J Syst Bacteriol. 1984;34:188–201. DOIGoogle Scholar
- Bousfield IJ, Green PN. Reclassification of bacteria of the genus Protomonas Urakami and Komagata 1984 in the genus Methylobacterium (Patt, Cole, and Hanson) emend. Green and Bousfield 1983. Int J Syst Bacteriol. 1985;35:209–10. DOIGoogle Scholar
- Hiraishi A, Furuhata K, Matsumoto A, Koike KA, Fukuyama M, Tabuchi K. Phenotypic and genetic diversity of chlorine-resistant Methylobacterium strains isolated from various environments. Appl Environ Microbiol. 1995;61:2099–107.PubMedGoogle Scholar
- La Scola B, Mallet MN, Grimont PAD, Raoult D. Bosea eneae sp. nov., Bosea massiliensis sp. nov. and Bosea vestrisii sp. nov. isolated from hospital water supplies, and emendation of the genus Bosea (Das et al.1996). Int J Syst Bacteriol. 2003;53:15–20.PubMedGoogle Scholar
- Bouzar H, Jones JB. Agrobacterium larrymoorei sp. nov., a pathogen isolated from aerial tumours of Ficus benjamina. Int J Syst Evol Microbiol. 2001;51:1023–6. DOIPubMedGoogle Scholar
- Bouzar H, Chilton WS, Nesme X, Dessaux Y, Vaudequin V, Petit A, A new Agrobacterium strain isolated from aerial tumors on Ficus benjamina L. Appl Environ Microbiol. 1995;61:65–73.PubMedGoogle Scholar
- Yabuuchi E, Kawamura Y, Kosako Y, Ezaki T. Emendation of genus Achromobacter and Achromobacter xylosoxidans (Yabuuchi and Yano) and proposal of Achromobacter ruhlandii (Packer and Vishniac) comb. nov., Achromobacter piechaudii (Kiredjian et al.) comb. nov., and Achromobacter xylosoxidans subsp. denitrificans (Ruger and Tan) comb. nov. Microbiol Immunol. 1998;42:429–38.PubMedGoogle Scholar
- Mudarris M, Austin B, Segers P, Vancanneyt M, Hoste B, Bernardet JF. Flavobacterium scophthalmum sp. nov., a pathogen of turbot (Scophthalmus maximus L.). Int J Syst Bacteriol. 1994;44:447–53. DOIPubMedGoogle Scholar
- Kumar S, Tamura K, Jakobsen IB, Nei M. MEGA2: Molecular evolutionary genetics analysis software. Bioinformatics. 2001;17:1244–5. DOIPubMedGoogle Scholar
- Drancourt M, Bollet C, Carlioz A, Martelin R, Gayral JP, Raoult D. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J Clin Microbiol. 2000;38:3623–30.PubMedGoogle Scholar
- Young JM, Kuykendall LD, Martinez-Romero E, Kerr A, Sawada H. A revision of Rhizobium Frank 1889, with an emended description of the genus, and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R. rubi, R. undicola and R. vitis. Int J Syst Evol Microbiol. 2001;51:89–103.PubMedGoogle Scholar
- Coenye T, Goris J, Spilker T, Vandamme P, Lipuma JJ. Characterization of unusual bacteria isolated from respiratory secretions of cystic fibrosis patients and description of Inquilinus limosus gen. nov., sp. nov. J Clin Microbiol. 2002;40:2062–9. DOIPubMedGoogle Scholar
- Birtles RJ, Rowbotham TJ, Raoult D, Harrison TG. Phylogenetic diversity of intra-amoebal legionellae as revealed by 16S rRNA gene sequence comparison. Microbiology. 1996;142:3525–30. DOIPubMedGoogle Scholar
- La Scola B, Mezi L, Auffray JP, Berland Y, Raoult D. Patients in the intensive care unit are exposed to amoeba-associated pathogens. Infect Control Hosp Epidemiol. 2002;23:462–5. DOIPubMedGoogle Scholar
- Fritsche TR, Horn M, Seyedirashti S, Gautom RK, Schleifer KH, Wagner M. In situ detection of novel bacterial endosymbionts of Acanthamoeba spp. phylogenetically related to members of the order Rickettsiales. Appl Environ Microbiol. 1999;65:206–12.PubMedGoogle Scholar
- Ly TM, Muller HE. Ingested Listeria monocytogenes survive and multiply in protozoa. J Med Microbiol. 1990;33:51–4. DOIPubMedGoogle Scholar
- Steinert M, Birkness K, White E, Fields B, Quinn F. Mycobacterium avium bacilli grow saprozoically in coculture with Acanthamoeba polyphaga and survive within cyst walls. Appl Environ Microbiol. 1998;64:2256–61.PubMedGoogle Scholar
- Horn M, Fritsche TR, Linner T, Gautom RK, Harzenetter MD, Wagner M. Obligate bacterial endosymbionts of Acanthamoeba spp. related to the beta-Proteobacteria: proposal of ‘Candidatus Procabacter acanthamoebae’ gen. nov., sp. nov. Int J Syst Evol Microbiol. 2002;52:599–605.PubMedGoogle Scholar
- Horn M, Harzenetter MD, Linner T, Schmid EN, Muller KD, Michel R, Members of the Cytophaga-Flavobacterium-Bacteroides phylum as intracellular bacteria of acanthamoebae: proposal of ‘Candidatus Amoebophilus asiaticus’. Environ Microbiol. 2001;3:440–9. DOIPubMedGoogle Scholar
- Rowbotham TJ. Isolation of Legionella pneumophila serogroup 1 from human feces with use of amebic cocultures. Clin Infect Dis. 1998;26:502–3. DOIPubMedGoogle Scholar
- Greub G, Berger P, Papazian L, Raoult D. Parachlamydiaceae as rare agents of pneumonia. Emerg Infect Dis. 2003;9:755–6.PubMedGoogle Scholar
Figures
Table
Cite This Article1G. Greub’s current affiliation is University of Lausanne, Lausanne, Switzerland.
Table of Contents – Volume 10, Number 3—March 2004
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Gilbert Greub, Microbiology institute IMU 02-219, Faculty of Biology and Medicine, University of Lausanne, Bugnon 48, 1011 Lausanne, Switzerland; fax: 00-41-21-314-40-60
Top