Volume 12, Number 6—June 2006
Dispatch
VIM-1 Metallo-β-lactamase in Acinetobacter baumannii
Abstract
In 2004 and 2005, 5 metallo-β-lactamase (MBL)-positive Acinetobacter baumannii isolates were found in 2 Greek hospitals. Isolates were unrelated and carried blaVIM-1 in a class 1 integron; blaOXA-51- and blaOXA-58-like carbapenemase genes were also detected. VIM-1 MBL in Acinetobacter spp. causes concern, given the increasing resistance of this species.
In the last few years, resistance to antibacterial drugs has been increasing in Acinetobacter spp., which will likely become a substantial treatment challenge in the future (1). Carbapenems have potent activity against Acinetobacter spp. and are usually the drugs of choice against multidrug-resistant Acinetobacter baumannii isolates. Acinetobacter spp. may develop resistance to carbapenems through various mechanisms, including class B and D carbapenemase production, decreased permeability, altered penicillin-binding proteins, and rarely, overexpression of efflux pumps (2,3).
In Europe, carbapenem resistance in A. baumannii has been sporadically attributed to the production of IMP-type metallo-β-lactamases (MBLs) and OXA-type carbapenemases (4). VIM-2–producing Acinetobacter spp. have been isolated in the Far East (4,5) and on 1 occasion in Germany (6). In this study, we report the appearance of the VIM-1 MBL determinant among A. baumannii in Greece.
We included in the study A. baumannii clinical isolates from tertiary care hospitals in 2 different Greek regions (Hippokration University Hospital, Thessaloniki, and University Hospital of Larissa, Thessalia) that were positive by the imipenem-EDTA double-disk synergy test (DDST) from March 2004 to March 2005. Bacteria were provisionally identified to the genus level by the Vitek 2 automated system (bioMérieux, Marcy l'Étoile, France) and the ATB 32GN system (bioMérieux). Antimicrobial drug susceptibility testing of the DDST-positive isolates for β-lactams (aztreonam, ceftazidime, cefepime, imipenem, meropenem, and piperacillin), β-lactam/β-lactamase inhibitor combinations (ampicillin/sulbactam, piperacillin/tazobactam), aminoglycosides (amikacin, gentamicin, netilmicin, and tobramycin), fluoroquinolones (ciprofloxacin and ofloxacin), and colistin was performed by Etest and Etest MBL (AB Biodisk, Solna, Sweden). The Clinical and Laboratory Standards Institute (CLSI) interpretative criteria were used (7), and Pseudomonas aeruginosa ATCC 27853 was used as control.
Polymerase chain reaction (PCR) testing of the synergy-positive isolates for carbapenemase genes was done by using consensus primers for blaIMP (8), blaVIM (9), blaSPM (10), blaOXA-23-like (11), blaOXA-24-like (11), blaOXA-58-like (11), and blaOXA-51-like (12). Pulsed-field gel electrophoresis (PFGE) of ApaI-digested genomic DNA was performed in the blaVIM-1-positive isolates, and the banding patterns were compared by using criteria proposed by Tenover et al. (13). The potential for conjugational transfer of imipenem resistance was examined in filter matings by using Escherichia coli 20R764 (lac+ rifr) as the recipient. Donor and recipient were mixed in a 1:5 ratio, and transconjugants were selected on MacConkey agar plates containing 100 μg/mL rifampicin and imipenem at concentrations of 0.5 to 2 μg/mL or 2 μg/mL ceftazidime.
Five A. baumannii clinical isolates that were MBL producers on the basis of DDST were detected among collections of isolates from patients hospitalized during the study period. Two of the isolates were recovered from blood cultures, 1 from bronchial secretions, 1 from a urine specimen, and 1 from cerebrospinal fluid; 2 isolates that had reduced susceptibility to carbapenems were not positive by the Etest MBL (Table). Imipenem MICs ranged from 4 to >32 μg/mL, while meropenem MICs were 2–32 μg/mL. P. aeruginosa ATCC 27853 was consistently characterized as having imipenem MIC of 2 μg/mL and meropenem MIC of 0.5 μg/mL. The 5 A. baumannii isolates were multidrug resistant; they showed resistance to all other antimicrobial drugs tested, with the exception of colistin.
Figure
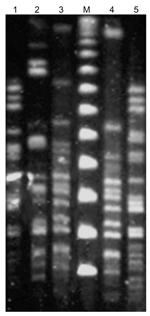
Figure. Pulsed-field gel electrophoresis of ApaI-restricted genomic DNA of the 5 blaVIM-1-positive Acinetobacter baumannii isolates. The index numbers of the isolates are those listed in the Table. Lane M, molecular mass marker (48.5 kb).
We did not detect blaIMP, blaSPM, blaOXA-23-like, or blaOXA-24-like in any of the 5 isolates, whereas blaVIM was detected in all of them. In 2 isolates, blaOXA-51- and blaOXA-58-like genes were also simultaneously present, while 2 more carried a blaOXA-51-like gene (Table). By sequencing both strands of the entire blaVIM amplicons (14), a blaVIM-1 sequence identical to that available in the database was identified. Sequencing blaOXA-51-like amplicons identified blaOXA-66 in all cases, while blaOXA-58-like alleles were classical blaOXA-58 in both cases. In 2 isolates, 1 from each region, PCR mapping of the integron that possibly carried blaVIM-1, with primers 5´ CS and a set of primers for genes aacA, dhfrI, aadA, qacEΔ1, and sul, showed a class 1 integron with a variable region including from 5´ to 3´ blaVIM-1, aacA7, dhfrI, and aadA1 gene cassettes. Sequencing of the overlapping PCR amplicons showed that this class 1 integron contained the intI1 gene with a strong P1 promoter, an inactivated (without a GGG insertion) P2 promoter, an attI1 site, and the blaVIM-1 gene cassette with its 59-base element identical to those reported previously in other gram-negative bacteria from Greece (15). PFGE showed that the 5 blaVIM-1-positive isolates did not form a genetically homogeneous group; they belonged to 4 distinct types. The 2 isolates from Thessaloniki were subtypes of the same clone (Table, Figure). In none of the 5 isolates was blaVIM-1 transferable to the susceptible E. coli host after repeated conjugal experiments with imipenem or ceftazidime selection. The 5,387-bp nucleotide sequence of the integron structure reported in this study has been submitted to the EMBL/GenBank/DDBJ sequence databases and has been assigned accession no. DQ112355.
This is the first report of the VIM-1 determinant in A. baumannii in the world. Occurrence of VIM-2 MBL among A. baumannii and Acinetobacter genomospecies 3 isolates has previously been described among clinical isolates in Korea (4,5). In Europe, IMP-type enzymes had been reported in single A. baumannii isolates from some European regions (4) and VIM-2 from 1 German hospital (6), while MBLs have not been detected in A. baumannii from the United States despite carbapenem resistance there (1). Though anticipated because of the circulation of blaVIM genes in several other gram-negative species in Europe and the ability of Acinetobacter spp. to acquire foreign DNA, this evolution is worrisome.
Retention of moderate susceptibility to carbapenems by blaVIM-positive A. baumannii isolates in our study may seem unexpected, since MBLs hydrolyze these compounds. However, MBL production in gram-negative bacteria may not substantially increase carbapenem MICs without the simultaneous operation of other mechanisms, such as impaired permeability (2,4). Furthermore, 2 of the 4 strains carrying oxacillinase genes with carbapenemase properties had imipenem MICs not higher than the CLSI breakpoints for resistance. Recently, blaOXA-51-like genes have been shown to be possibly naturally occurring (12), while OXA-58 enzymes play a minor role in carbapenem resistance in A. baumannii, and strong promoter sequences are needed for higher levels of resistance to carbapenems (2).
During the last few years, A. baumannii has been increasingly isolated from severely ill patients, and its usual cross-resistance to most available antimicrobial drugs, including carbapenems, poses substantial problems worldwide and especially in the United States (1). In New York approximately two thirds of isolates are carbapenem resistant (3). In our region, Acinetobacter spp. are frequent nosocomial pathogens and are commonly multidrug resistant, which leads to the extensive use of carbapenems and, lately, polymyxins. The presence of MBLs among carbapenem-resistant Acinetobacter spp. from different Greek regions emphasizes the need for restricted use of carbapenems and early recognition of strains producing these enzymes. Although Etest MBL was reliable to detect VIM-2-producing Acinetobacter spp. in Korea (5), the assay seems unable to identify MBL-positive isolates exhibiting relatively low carbapenem MICs. Therefore, our diagnostic laboratories should screen Acinetobacter spp. with imipenem-EDTA DDST or alternative DDSTs, such as those using 2-mercaptopropionic acid, which appears to be more sensitive for detecting MBLs among Acinetobacter spp (4). Whether carbapenems might be appropriate to treat infections with low-level carbapenem-resistant or susceptible blaVIM-bearing A. baumannii isolates has yet to be determined.
Dr Tsakris is associate professor of microbiology at the University of Athens, Greece. His research interests include the investigation of antimicrobial drug resistance mechanisms of gram-negative pathogens and the molecular epidemiology of hospital infections.
References
- Gaynes R, Edwards JR; National Nosocomial Infections Surveillance System. Overview of nosocomial infections caused by gram-negative bacilli. Clin Infect Dis. 2005;41:848–54. DOIPubMedGoogle Scholar
- Heritier C, Poirel L, Lambert T, Nordmann P. Contribution of acquired carbapenem-hydrolyzing oxacillinases to carbapenem resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49:3198–202. DOIPubMedGoogle Scholar
- Quale J, Bratu S, Landman D, Heddurshetti R. Molecular epidemiology and mechanisms of carbapenem resistance in Acinetobacter baumannii endemic in New York City. Clin Infect Dis. 2003;37:214–20. DOIPubMedGoogle Scholar
- Walsh TR, Toleman MA, Poirel L, Nordmann P. Metallo-β-lactamases: the quiet before the storm? Clin Microbiol Rev. 2005;18:306–25. DOIPubMedGoogle Scholar
- Lee K, Lee WG, Uh U, Ha GY, Cho J, Chong Y, VIM- and IMP-type metallo-β-lactamase-producing Pseudomonas spp. and Acinetobacter spp. in Korean hospitals. Emerg Infect Dis. 2003;9:868–71.PubMedGoogle Scholar
- Toleman MA, Jones RN, Walsh TR. Hospital outbreak of an imipenem-resistant VIM-2 encoding Acinetobacter DNA group 14TU strain in a German teaching hospital. Clin Microbiol Infect. 2004;10(Suppl 3):48.PubMedGoogle Scholar
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing; 15th informational supplement. CLSI/NCCLS M100-S15. Wayne (PA): The Institute; 2005.
- Senda K, Arakawa Y, Ichiyama S, Nakashima K, Ito H, Ohsuka S, PCR detection of metallo-β-lactamase gene (blaIMP) in gram-negative rods resistant to broad-spectrum β-lactams. J Clin Microbiol. 1996;34:2909–13.PubMedGoogle Scholar
- Tsakris A, Pournaras S, Woodford N, Palepou MFI, Babini GS, Douboyas J, Outbreak of infections caused by Pseudomonas aeruginosa producing VIM-1 carbapenemase in Greece. J Clin Microbiol. 2000;38:1290–2.PubMedGoogle Scholar
- Toleman MA, Simm AM, Murphy TA, Gales AC, Biedenbach DJ, Jones RN, Molecular characterization of SPM-1, a novel metallo-β-lactamase isolated in Latin America: report from the SENTRY antimicrobial surveillance programme. J Antimicrob Chemother. 2002;50:673–9. DOIPubMedGoogle Scholar
- Poirel L, Marqué S, Héritier C, Segonds C, Chabanon G, Nordmann P. OXA-58, a novel class D β-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49:202–8. DOIPubMedGoogle Scholar
- Héritier C, Poirel L, Fournier PE, Claverie JM, Raoult D, Nordmann P. Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49:4174–9. DOIPubMedGoogle Scholar
- Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233–9.PubMedGoogle Scholar
- Yan JJ, Hsueh PR, Ko WC, Luh KT, Tsai SH, Wu HM. Metallo-β-lactamases in clinical Pseudomonas isolates in Taiwan and identification of VIM-3, a novel variant of the VIM-2 enzyme. Antimicrob Agents Chemother. 2001;45:2224–8. DOIPubMedGoogle Scholar
- Giakkoupi P, Xanthaki A, Kanelopoulou M, Vlahaki A, Miriagou V, Kontou S, VIM-1 metallo-β-lactamase-producing Klebsiella pneumoniae strains in Greek hospitals. J Clin Microbiol. 2003;41:3893–6. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleTable of Contents – Volume 12, Number 6—June 2006
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Athanassios Tsakris, Department of Microbiology, Medical School, University of Athens, 75 Mikras Asias St, 115 27 Athens, Greece
Top