Volume 14, Number 6—June 2008
Letter
Mycobacterium avium subsp. hominissuis Infection in 2 Pet Dogs, Germany
Cite This Article
Citation for Media
To the Editor: The genus Mycobacterium contains various obligate and opportunistic pathogens of animals, which may also be transmitted to humans and cause disease in, thus exhibiting a considerable zoonotic potential (1,2). During the past few decades, members of the Mycobacterium avium-intracellulare complex (MAIC) emerged as pathogens of human diseases, including lymphadenitis in children, pulmonary tuberculosis-like disease, and disseminated infections (occurring predominantly in immunocompromised persons, particularly AIDS patients) (1,2). Similarly, important animal diseases are caused by members of this group, e.g., avian tuberculosis and paratuberculosis in ruminants (1). MAIC includes M. intracellulare and 4 subspecies of M. avium, namely, M. avium subsp. avium, M. avium subsp. hominissuis, M. avium subsp. silvaticum, and M. avium subsp. paratuberculosis (3,4). Whereas members of the M. tuberculosis complex are transmitted by direct host contact, MAIC species are acquired predominantly from environmental sources, including soil, water, dust, and feed. Subclinical infections are common among birds (1,2).
M. avium strains differ from M. intracellulare by containing the insertion sequence (IS) IS1245 (3) and are further discriminated by terms of IS901 (4). Avian isolates (M. avium subsp. avium) are usually positive for IS901 and represent the main pathogen of avian tuberculosis (5). In contrast, mammalian isolates are IS901-negative and have been designated as M. avium subsp. hominissuis because of their predominant hosts. This subspecies is only weakly virulent for birds but causes disease in animals and humans (5).
Even though M. tuberculosis and M. bovis are the common etiologic agents of canine mycobacteriosis, dogs are reported to be relatively resistant to M. avium infection (6,7). Nonetheless, sporadic cases usually show nonspecific clinical signs, whereas necropsy consistently reveals granulomatous inflammation in numerous organs, including lymph nodes, intestine, spleen, liver, lung, bone marrow, and even spinal cord (7,8). The predominant involvement of the gastrointestinal tract indicates an oral route of infection (7,8), and simultaneously increases the risk for human infection by fecal spread of mycobacteria.
Our report concerns 2 young dogs, a 3-year-old miniature schnauzer and a 1-year-old Yorkshire terrier, that lived in different geographic regions in Germany. Both had had therapy-resistant fever, lethargy, progressive weight loss, and generalized lymphadenomegaly for several weeks and were euthanized after a final phase of diarrhea. Necropsy findings, similar in both dogs, included generalized enlargement of lymph nodes with a whitish, granular to greasy cut surface, leading to intraabdominal adhesions by extensive involvement of mesenteric lymph nodes. In the terrier, the greater omentum and a part of the right apical lung lobe showed changes similar to those in the lymph nodes. Furthermore, numerous white 1-mm nodules were found in the spleen (both dogs), liver (schnauzer) and costal pleura (terrier).
Figure
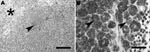
Figure. A) Mesenteric lymph node of Yorkshire Terrier shows diffuse granulomatous lymphadenitis with extensive infiltration of macrophages, foci of pyogranulomatous inflammation (arrowhead), and focal necrosis (asterisk). Hematoxylin and eosin stain; scale bar...
Histologic examination showed (pyo-)granulomatous inflammation of lymph nodes, tonsils, liver, spleen, and greater omentum. Additionally, pyogranulomatous pleuropneumonia was present in the terrier, and a granulomatous enteritis and pyelitis in the schnauzer. The granulomatous lesions frequently exhibited central necrosis surrounded by macrophages, epitheloid cells, and few neutrophils (Figure, panel A). However, multinucleated giant cells or mineralization was not observed. In both animals, Ziehl-Neelsen stain demonstrated large numbers of acid-fast bacilli within macrophages (Figure, panel B). Samples of lymph nodes and lung were processed for mycobacterial culture by using standard procedures (Löwenstein-Jensen, Stonebrink medium). Colonies emerging after 2-week incubation at 37°C were investigated by PCR targeting IS1245 and IS901 (3,4). In all samples, M. avium subsp. hominissuis was identified by growth characteristics as well as presence of an IS1245-specific and absence of an IS901-specific PCR product. Additionally, sequencing of hsp65 was conducted (9), which indicated M. avium subsp. hominissuis in both dogs (GenBank accession nos. EU488724 and EU488725).
Despite improved therapeutic approaches, MAIC infection represents a frequent bacterial complication in persons with AIDS. However, several studies showed a very low incidence of M. avium subsp. avium infections in humans. Thus, most of these HIV-related infections are attributed to M. avium subsp. hominissuis (2,5). Unfortunately, the subspecies of M. avium was not identified in most canine cases reported in the literature (7,8). Nonetheless, different serotypes of M. avium, corresponding to either M. avium subsp. avium or M. avium subsp. hominissuis, have been identified sporadically (6,10). The source and route of infection were unclear in all reports including ours, albeit repeatedly observed enteritis strongly suggested an oral mode of infection. A common environmental or wildlife reservoir represents the most probable source of M. avium infection for both humans and animals. However, there is also evidence of direct transmission (1–3). Therefore, M. avium subsp. hominissuis infection in dogs may comprise a considerable zoonotic potential, particularly if pet dogs with close contact to the owner are affected and if prolonged nonspecific clinical signs and intestinal involvement occur, as demonstrated here.
Acknowledgment
We thank Bettina Buck, Heidrun Wittig, Sandy Werner, and Karola Zmuda for their excellent technical assistance.
References
- Biet F, Boschiroli ML, Thorel MF, Guilloteau LA. Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium–intracellulare complex (MAC). Vet Res. 2005;36:411–36. DOIPubMedGoogle Scholar
- Ashford DA, Whitney E, Raghunathan P, Cosivi O. Epidemiology of selected mycobacteria that infect humans and other animals. Rev Sci Tech. 2001;20:325–37.PubMedGoogle Scholar
- Guerrero C, Bernasconi C, Burki D, Bodmer T, Telenti A. A novel insertion element from Mycobacterium avium, IS1245, is a specific target for analysis of strain relatedness. J Clin Microbiol. 1995;33:304–7.PubMedGoogle Scholar
- Kunze ZM, Portaels F, McFadden JJ. Biologically distinct subtypes of Mycobacterium avium differ in possession of insertion sequence IS901. J Clin Microbiol. 1992;30:2366–72.PubMedGoogle Scholar
- Mijs W, de Haas P, Rossau R, Van der Laan T, Rigouts L, Portaels F, Molecular evidence to support a proposal to reserve the designation Mycobacterium avium subsp. avium for bird-type isolates and ‘M. avium subsp. hominissuis’ for the human/porcine type of M. avium. Int J Syst Evol Microbiol. 2002;52:1505–18. DOIPubMedGoogle Scholar
- Friend SC, Russell EG, Hartley WJ, Everist P. Infection of a dog with Mycobacterium avium serotype II. Vet Pathol. 1979;16:381–4.PubMedGoogle Scholar
- O’Toole D, Tharp S, Thomsen BV, Tan E, Payeur JB. Fatal mycobacteriosis with hepatosplenomegaly in a young dog due to Mycobacterium avium. J Vet Diagn Invest. 2005;17:200–4.PubMedGoogle Scholar
- Horn B, Forshaw D, Cousins D, Irwin PJ. Disseminated Mycobacterium avium infection in a dog with chronic diarrhoea. Aust Vet J. 2000;78:320–5. DOIPubMedGoogle Scholar
- Carpenter JL, Myers AM, Conner MW, Schelling SH, Kennedy FA, Reimann KA. Tuberculosis in five basset hounds. J Am Vet Med Assoc. 1988;192:1563–8.PubMedGoogle Scholar
- Turenne CY, Semret M, Cousins DV, Collins DM, Behr MA. Sequencing of hsp65 distinguishes among subsets of the Mycobacterium avium complex. J Clin Microbiol. 2006;44:433–40. DOIPubMedGoogle Scholar
Figure
Cite This ArticleRelated Links
Table of Contents – Volume 14, Number 6—June 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Peter Wohlsein, Department of Pathology, University of Veterinary Medicine Hanover, Buenteweg 17, D-30559 Hanover, Germany;
Top