Volume 21, Number 9—September 2015
Dispatch
Putative Lineage of Novel African Usutu Virus, Central Europe
Cite This Article
Citation for Media
Abstract
We characterized the complete genome of a putative novel Usutu virus (USUV) strain (Usutu-BONN) detected in a dead blackbird from Germany. Genomic analysis revealed several unique amino acid substitutions among the polyprotein gene. Phylogenetic analyses demonstrated that Usutu-BONN constitutes a putative novel African USUV lineage, which was probably recently introduced to central Europe.
Originally isolated from a Culex neavei mosquito in South Africa in 1959 (1,2), Usutu virus (USUV) was subsequently detected in different mosquito and bird species throughout Sub-Saharan countries (3). USUV has recently been introduced to Europe, where it caused widespread deaths among resident bird populations, established a local transmission cycle, and became a resident pathogen (4–6). USUV is maintained in an enzootic cycle involving mosquitoes as vectors and birds as the main amplifying hosts; humans are considered incidental or dead-end hosts. We have demonstrated that bats could also be infected with USUV and might act as amplifying hosts (7), and there is increasing evidence that USUV is pathogenic for humans, thus becoming a potential public health problem (8,9). On the basis of genetic differences, in comparison with the USUV strains from Africa, the USUV strains from Europe, except those from Spain, form a distinct clade within USUV phylogeny (7). The detection and isolation of USUV from different bird species and mammalophilic mosquitoes during the 2011 epizootic in Germany raised questions regarding the USUV host range. Thus, as a part of the German Arbovirus Surveillance Program (10), we continued the monitoring of the mosquitoes, birds, and bats for the presence of USUV.
Figure 1
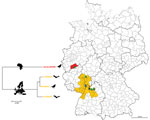
Figure 1. Region of Germany where Usutu virus (USUV) is endemic (orange) and location where the putative novel USUV strain Usutu-BONN was detected (red). Phylogenetic tree illustrates the genetic relationship between the strains...
During May–October 2014, ≈23,300 female mosquitoes from different parts of Germany were trapped, morphologically identified, and pooled (up to 250 mosquitoes/pool). During January–November 2014, a total of 8 dead Pipistrellus bats and 32 dead birds (mainly blackbirds) from different regions of the country were subjected to complete necropsy, and samples were collected for virus detection. Total RNA and DNA from homogenized mosquito pools and tissue samples (brain, liver, lung, and heart) from bats and birds were extracted by using an RTP DNA/RNA Virus Mini Kit (STRATEC Biomedical, Birkenfeld, Germany) according the manufacturer’s instructions. Extracted samples were analyzed for the presence of flavivirus RNA by using a modified pan-flavivirus reverse transcription PCR (5). Positive results were found for 5 mosquito pools (C. pipiens biotype pipiens), 5 blackbirds (Turdus merula), and 1 bat (Pipistrellus pipistrellus). Direct sequencing of the pan-flavivirus PCR amplicons showed USUV nucleic acid sequences in each sample. The positive samples were further subjected to PCRs for the amplification of a partial segment of USUV envelope and nonstructural (NS) 5 gene. Sequencing results showed that all samples, except 1 blackbird-derived USUV sequence, were identical (data not shown) and originated from southwest Germany, corresponding to the previously described USUV-endemic area (Figure 1) (5,10,11). The USUV-positive blackbird sample, which exhibited numerous nucleotide and amino acid changes compared with other sequences, had been found outside of the USUV-endemic area, at the beginning of August in Sankt Augustin (50°46′12′′N, 7°11′12′′E), a city located near Bonn.
Figure 2

Figure 2. Phylogenetic relationship of the Usutu virus Usutu-BONN strain (from Germany, 2014) and other Usutu viruses, based on complete amino acid sequences of the polyprotein-encoding gene. The phylogenetic tree was constructed by...
Full-length genome sequence of this putative novel USUV strain, designated Usutu-BONN (GenBank accession no. KM659876), was successfully obtained by using a previously described protocol (7). The genome contained 11,065 nt with a 96-nt 5′ untranslated region and a 664-nt 3′ untranslated region. The single open reading frame encoded a polyprotein of 3,434 aa. Depending on the USUV strain, the nucleotide sequence similarity ranged from 81% to 98%, whereas amino acid sequence conservation ranged from 94.7% to 99.2% (Table 1). Comparison of the Usutu-BONN complete polyprotein sequence with the other USUV strains showed several identical synonymous and nonsynonymous mutations characteristic for African USUV strains and some unique substitutions (Table 2). Usutu-BONN contained 15 aa substitutions, 7 of which were unique mutations (Table 2). Most were located in the envelope (E) and NS2a proteins (Table 2). We also found 1 putative change in the C/anchC cleavage site of the polyprotein (Table 2); this mutation was observed also in the highly divergent African USUV strain (ArB1803; GenBank accession no. KC754958). We detected 2 more unique amino acid substitutions in the domain II of the E protein. Strikingly, no potential N-glycosylation sites or substitutions were found in well-known features of the protein E fusion peptide or antibody binding sites. Unique mutations (V142 and V189) in the NS2a protein were also observed (Table 2). Alignment of the deduced amino acid sequences of the complete polyprotein (including all previously described strains for which complete polyprotein-encoding sequences are available) by using the MAFFT plugin in Geneious 7.1.5 (Biomatters, Ltd, Auckland, New Zealand) and subsequent phylogenetic reconstruction by using a maximum-likelihood tree (JTT+G model) with 1,000 bootstrap replicates in PhyML (12) and parallel Bayesian Markov Chain Monte Carlo method implemented in MrBayes 3.0 software (13) (data not shown) demonstrated that Usutu-BONN forms a separate lineage (basal position of the African clade) within the USUV phylogeny (Figure 2). Furthermore, the phylogenetic tree showed moderate genetic relatedness of the Usutu-BONN strain with USUV strains circulating in central Europe.
We detected and genetically characterized a putative novel USUV strain (Usutu-BONN) by determining its complete genome sequence and comparing it with USUV strains for which complete polyprotein-encoding sequences are available. We demonstrated that the Usutu-BONN strain from Germany constitutes a putative novel USUV lineage. The unique synonymous mutations detected in the E and NS2a genes of Usutu-BONN strains may suggest an adaptive evolution. In this strain, 1 putative cleavage site of the viral polyprotein responsible for processing of structural proteins was changed. Given that Usutu-BONN has not led to massive deaths among birds and has not yet been found in other hosts or mosquito vectors, it seems evident that this strain was recently introduced into Germany and evolved in another geographic region, probably Africa (Figure 1, 2). The possibility of an African origin of this virus strain is strengthened by the fact that phylogenetic analysis of complete polyprotein sequence established a separate basal lineage for the Usutu-BONN strain in a sister relationship with the African USUV strains, suggesting that Usutu-BONN has evolved in parallel with strains from Africa sharing a recent common ancestor. This putative novel USUV strain was introduced into Europe probably as other strains, via viremic migratory birds returning from winter migration from Africa to Europe or through ship- or aircraft-borne transportation of USUV-infected mosquitoes from Africa. However, identification of the possible sources (e.g., infected mosquitoes, resident or short-ranging migratory birds) of this new USUV strain will require sequence information from neighboring countries where USUV has been detected. The detection of USUV in a Pipistrellus bat 1 year after the first detection of USUV in bats from the same area in 2013 further strengthens our previous hypothesis that bats may contribute to the epizootic by serving as amplifying/reservoir hosts (7). The unique mutations (V142 and V189) in the NS2a protein of the Usutu-BONN strain are located very close to the identical mutation observed in the previously described bat-derived USUV strains. Although the biological consequences of these mutations are not known, similar mutations in West Nile virus were responsible for inhibition of interferon signaling (14).
Further monitoring studies are necessary to evaluate the pathogenic potential of this newly introduced USUV strain in central Europe for susceptible and receptive avian/mammalian hosts and for humans. This information could be used to predict future epidemics and to implement adequate preventive and control measures.
Dr. Cadar is a postdoctoral researcher at the Bernhard Nocht Institute for Tropical Medicine, Hamburg, Germany. His research interests include the arbovirus ecology, evolutionary genetics of viral emergence, and arbovirus discovery.
Acknowledgment
We thank Alexandra Bialonski, Martin Rudolf, Claudia Poggensee, Mathis Petersen, Birgit Hüsing, Corinna Thomè, and Sabine Köhler for excellent technical assistance.
References
- Williams MC, Simpson DI, Haddow AJ, Knight EM. The isolation of West Nile virus from man and of Usutu virus from the bird-biting mosquito Mansonia aurites (Theobald) in the Entebbe area of Uganda. Ann Trop Med Parasitol. 1964;58:367–74 .PubMedGoogle Scholar
- McIntosh BM. Usutu (SA Ar 1776), nouvel arbovirus du groupe B. International Catalogue of Arboviruses. 1985;3:1059–60.
- Nikolay B, Diallo M, Boye CS, Sall AA. Usutu virus in Africa. Vector Borne Zoonotic Dis. 2011;11:1417–23 . DOIPubMedGoogle Scholar
- Weissenböck H, Kolodziejek J, Url A, Lussy H, Rebel-Bauder B, Nowotny N. Emergence of Usutu virus, an African mosquitoborne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg Infect Dis. 2002;8:652–6. DOIPubMedGoogle Scholar
- Becker N, Jöst H, Ziegler U, Eiden M, Höper D, Emmerich P, Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE. 2012;7:e32604. http://dx.doi.org/10.1371/ journal.pone.0032604.
- Savini G, Monaco F, Terregino C, Di Gennaro A, Bano L, Pinoni C, Usutu virus in Italy: an emergence or a silent infection? Vet Microbiol. 2011;151:264–74. DOIPubMedGoogle Scholar
- Cadar D, Becker N, Campos Rde M, Börstler J, Jöst H, Schmidt-Chanasit J. Usutu virus in bats, Germany, 2013. Emerg Infect Dis. 2014;20:1771–3. DOIPubMedGoogle Scholar
- Pecorari M, Longo G, Gennari W, Grottola A, Sabbatini A, Tagliazucchi S, First human case of Usutu virus neuroinvasive infection, Italy, August–September 2009. Euro Surveill. 2009;14:19446 .PubMedGoogle Scholar
- Vilibic-Cavlek T, Kaic B, Barbic L, Pem-Novosel I, Slavic-Vrzic V, Lesnikar V, First evidence of simultaneous occurrence of West Nile virus and Usutu virus neuroinvasive disease in humans in Croatia during the 2013 outbreak. Infection. 2014;42:689–95. DOIPubMedGoogle Scholar
- Becker N, Krüger A, Kuhn C, Plenge-Bönig A, Thomas SM, Schmidt-Chanasit J, Mosquitoes as vectors for exotic pathogens in Germany. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz. 2014;57:531–40. DOIPubMedGoogle Scholar
- Jöst H, Bialonski A, Maus D, Sambri V, Eiden M, Groschup MH, Isolation of Usutu virus in Germany. Am J Trop Med Hyg. 2011;85:551–3. DOIPubMedGoogle Scholar
- Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol. 2010;59:307–21 .DOIPubMedGoogle Scholar
- Ronquist F, Huelsenbeck JP. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–4. DOIPubMedGoogle Scholar
- Liu WJ, Wang XJ, Mokhonov VV, Shi PY, Randall R, Khromykh AA. Inhibition of interferon signaling by the New York 99 strain and Kunjin subtype of West Nile virus involves blockage of STAT1 and STAT2 activation by nonstructural proteins. J Virol. 2005;79:1934–42 .DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 21, Number 9—September 2015
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Jonas Schmidt-Chanasit, Bernhard Nocht Institute for Tropical Medicine, WHO Collaborating Centre for Arbovirus and Hemorrhagic Fever Reference and Research, Bernhard-Nocht-Strasse 74, 20359 Hamburg, Germany
Top