Volume 5, Number 6—December 1999
Perspective
Emerging Infectious Diseases and Amphibian Population Declines
Chytridiomycosisan Emerging Panzootic Fungal Disease of Amphibians
Emergence of Chytridiomycosis
Chytridiomycosis as the Cause of Population Declines
Potential Environmental Cofactors in the Emergence of Chytridiomycosis
Emerging Viral Diseases of Amphibians
Characteristics of Ranaviral Disease
Factors Associated with the Emergence of Ranaviruses
Impact of Ranaviral Disease on Amphibian Populations
Implications of Emerging Infectious Diseases for Amphibian Population Declines and Wildlife Conservation
Cite This Article
Cite This Article
Citation for Media
Abstract
We review recent research on the pathology, ecology, and biogeography of two emerging infectious wildlife diseases, chytridiomycosis and ranaviral disease, in the context of host-parasite population biology. We examine the role of these diseases in the global decline of amphibian populations and propose hypotheses for the origins and impact of these panzootics. Finally, we discuss emerging infectious diseases as a global threat to wildlife populations.
Emerging infectious diseases have been reported increasingly as causes of death in free-living wild animals (1). These diseases are a particular threat to wildlife species whose population, habitat, or range has been diminished or artificially manipulated to promote species survival (e.g., captive breeding, translocation, and release programs) (2-4). An early example of an emerging disease panzootic was the introduction of rinderpest in African domestic cattle in 1889 (5). More recently, epizootics and panzootics of wildlife have been increasingly reported in terrestrial (1) and marine (6) habitats and are probably underreported (1,4,7-9). Recent advances in theoretical and experimental host-parasite ecology have demonstrated a major role for infectious agents in the population biology of wild animals (10,11). We discuss recent data on two newly emerging infectious diseases of amphibians and, by reference to host-parasite ecology, propose hypotheses to explain their origin and impact.
Global declines in amphibian population are perhaps one of the most pressing and enigmatic environmental problems of the late 20th century (12-19). While some declines are clearly due to habitat destruction, others are not associated with obvious environmental factors. Causal hypotheses include the introduction of predators or competitors, increased ultraviolet (UV-B) irradiation, acid precipitation, adverse weather patterns, environmental pollution, infectious disease, or a combination of these. Transdermal water uptake and gaseous exchange and a biphasic life cycle are important aspects of amphibian biology. These factors led to the hypothesis that amphibians act as sentinels for global environmental degradation (12,18). However, this role has yet to be demonstrated, and many causal factors may be present (12,19,20).
Of particular concern are population declines in ecologically pristine areas, such as the montane tropical rain forests of Australia and Central America, where human impact from agriculture, deforestation, or pollution is thought to be negligible. Here, long-term data demonstrate recent and catastrophic amphibian population declines, often resulting in the complete loss of amphibian species (local extinction of multiple species) from large swaths of habitat (20-25). These declines include the disappearance and presumed extinction of the recently discovered golden toad (Bufo periglenes) of Costa Rica (23) and as many as seven Australian amphibian species, including two species of gastric-brooding frog (Rheobatrachus spp.) (20). These data, along with recent findings of amphibian mass deaths in these areas, suggest that such local extinctions are not normal population fluctuations or metapopulation dynamics.
Investigations in Australia, the United Kingdom, and North and Central America (26-31) have repeatedly found two diseases as the causes of amphibian mass deaths globally (Table 1): chytridiomycosis in the rain forests of Australia and Central and South America and some parts of North America (28-32) and iridoviral infections in the United Kingdom, the United States, and Canada (26,27,33-36). Both diseases have been classified as emerging (1). The parasitic infection recently implicated as the cause of amphibian deformities in North America has not been associated with mass deaths or population declines (31).
Figure 1
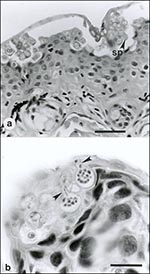
Figure 1. . A: Ventral abdominal skin of Bufo haematiticus from western Panama. The superficial keratinized layer of epidermis (stratum corneum) contains numerous intracellular spherical-to-ovoid sporangia (spore-containing bodies) of Batrachochytrium sp. The mature...
Figure 2
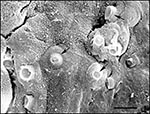
Figure 2. Scanning electron micrograph of digital skin of a wild frog (Litoria lesueuri, from Queensland, Australia) that died of cutaneous chytridiomycosis. Many cells within this area of the superficial layer of the...
Chytridiomycosis is a fungal disease first described in 1998 from moribund and dead adult amphibians collected at sites of mass deaths in Australia and Panama from 1993 to 1998 (28). Here, long-term ecologic study sites reported catastrophic amphibian population declines in Big Tableland, Queensland (39,40), and Fortuna and Cerro Pando, Panama (24,25,28). No significant pathogens were found on routine parasitologic, bacteriologic, mycologic, or virologic examinations of tissue samples (28). Fresh skin smears and histologic sections of the epidermis, however, consistently contained large numbers of developing and mature sporangia of a new genus of chytrid fungus (phylum Chytridiomycota) (Figures 1, 2). Sporangia were also present within the keratinized mouthparts, but not the epidermis, of sympatric tadpoles (tadpoles lack epidermal keratin) (28,29). No significant morphologic differences between chytrids infecting Australian and Central American amphibians were found by transmission electron microscopy, and the pathogen was identified as a member of the order Chytridiales by analysis of zoospore ultrastructure and 18s rDNA sequence data (28). Chytrids are ubiquitous fungi that develop without hyphae and are found in aquatic habitats and moist soil, where they degrade cellulose, chitin, and keratin (41). Parasitic chytrids mainly infect plants, algae, protists, and invertebrates (41); the amphibian pathogen is the only example of a chytrid parasitizing vertebrates (28).
Clinical signs of amphibian chytridiomycosis include abnormal posture, lethargy, and loss of righting reflex. Gross lesions, which are usually not apparent, consist of abnormal epidermal sloughing and (more rarely) epidermal ulceration; hemorrhages in the skin, muscle, or eye; hyperemia of digital and ventrum skin, and congestion of viscera (29). Diagnosis is by identification of characteristic intracellular flask-shaped sporangia and septate thalli within the epidermis (Figures 1, 2).
Its occurrence solely in keratinized tissue suggests that the chytrid uses amphibian keratin as a nutrient. A hyperkeratotic and hyperplastic response of the epidermis to infection (restricted to the stratum corneum and stratum granulosum) usually coincides with the immediate location of chytrid developmental stages. Inflammatory cell response is negligible. An isolate cultured from captive dendrobatid frogs has recently been used to fulfil Koch's postulates as a fatal pathogen of frogs and has been described as a new genus and species, Batrachochytrium dendrobatidis (42). Three mechanisms by which chytridiomycosis causes death have been proposed (28): 1) epidermal hyperplasia impairs essential cutaneous respiration or osmoregulation; 2) a fungal toxin is absorbed systemically (although a lack of clinical signs in infected larvae suggests otherwise); and 3) these factors are combined (28,37).
Comparison of histologic, ultrastructural, and 18s rDNA sequence data indicates that the chytrids found in wild Australian (28) and captive Central American (T.Y. James, D. Porter, J.E. Longcore, pers. comm.) amphibians belong to the genus Batrachochytrium, are probably conspecific, and form a distinct monophyletic clade within the Chytridiales.
Retrospective histologic surveys of museum specimens of montane, riparian anurans from protected sites in Central America and Australia, conducted 1 to 10 years before the population declines, showed no evidence of chytrid infection, which suggests that chytridiomycosis has recently emerged on two continents (28). The relatively synchronous discovery of chytridiomycosis in Australia and Central America in association with amphibian population declines is striking. The data suggest that Batrachochytrium 1) may be endemic to these regions and the amphibian deaths and declines attributed to it have only recently been discovered; 2) may be endemic and has recently become pathogenic (e.g., through an increase in the organism's prevalence or virulence, or a decrease in the host's defenses), or 3) may have been introduced recently into these geographic regions and is now parasitizing novel host species.
The pattern of amphibian deaths and population declines associated with chytridiomycosis is characteristic of an introduced virulent pathogen dispersing through a naïve population (7,10,39). In Australia, a distinct geographic and temporal progression in population declines has occurred (20), moving northward at a mean rate of 100 km per year (39). In Central America, a progression from northern Costa Rica to western Panama occurred from 1996 to 1998 (24,25). The uneven progression of declines in Australia (40) may reflect gaps in surveillance. Small-scale irregularities, however, characterize the epidemiology of many pandemics (43,44), within which individual epidemics progress at different rates in different areas. In amphibian populations, unevenness may be due to differences in ecologic factors (e.g., population density, habitat, age structure); differences among pathogen strains; stochastic factors, such as the time of introduction; or a combination of these factors. In Australia and Central America, population declines have been catastrophic, occurring over a few months, with dramatic population loss and high rates of adult deaths (20,24,25,28,39,40,45). Such high depopulation rates are characteristic of introduced virulent pathogens (10). Conversely, in coevolved host-pathogen relationships, a degree of herd immunity to the pathogen and lower virulence (infectivity and death rates) are normally observed. The low host specificity of amphibian chytridiomycosis (more than 30 species of wild amphibians from seven families in Central America and Australia [28,29]) also suggests that the disease was not enzootic in those montane rain-forest populations. Ability to infect a range of host species is a characteristic of many invading pathogens (10) and is less common in endemic microparasites that have coevolved with their hosts.
The most parsimonious hypothesis for the origin of chytridiomycosis panzootics in Australia and Central America is the introduction of disease into populations of previously unexposed amphibians. Introduction of pathogens, termed pathogen pollution (1), is increasingly recognized as a significant threat to global biodiversity (1,6,46) and forms an integral part of human history (10,47). There are precedents for the introduction of fungal pathogens (including chytrid parasites) that cause high death rates (41,48). Mechanisms by which wildlife pathogens can be introduced are common; for example, a consequence of the increasing mobility of humans is the global translocation of wildlife, plants, soil, and ballast water (1,4,49,50). Freshwater fish and amphibians are also transported globally. In Australia, chytridiomycosis-infected cane toads (Bufo marinus), a recently introduced species, have been found (29), and in North America, bullfrogs (Rana catesbeiana) and other species of amphibians have been translocated or introduced widely. Some authors have suggested that tourists or fieldworkers surveying amphibian populations may have facilitated the dissemination of Batrachochytrium (19), although this has not been demonstrated. Batrachochytrium may have coevolved with some amphibians (e.g., lowland) species, populations of which remain unaffected. Recent disturbances of rain forest habitats may have introduced this parasite into naïve populations in Central America and Australia, leading to mass deaths. This range of disease outcomes parallels many diseases of humans, e.g., measles and smallpox, which produce a range of effects on persons in disease-endemic regions and cause massive deaths when introduced into naïve populations (48).
The occurrence of chytridiomycosis in free-living North American amphibians (Table 1) suggests a less obvious pattern of dissemination than in Central America and Australia. This irregularity may be due to a paucity of data, the pathogen's being enzootic to the United States, or the pathogen's introduction a number of years before. Historical reports of declines in the United States include postmetamorphic death syndrome, which progressed in waves through populations of amphibians, causing 90% to 100% death rates in recently metamorphosed animals and low death rates in larvae (50). Recent reports of chytridiomycosis-linked die-offs in Bufo boreas markedly resemble these previous die-offs. This observation and the finding that chytridiomycosis caused similar die-offs (of B. canorus and R. pipiens) in the 1970s (Table 1) (32), support the last of the above hypotheses. In the United States an amphibian pathogen (histologically very similar to Batrachochytrium but identified as Basidiobolus ranarum) has been described in wild Wyoming toads (Bufo hemiophrys baxteri) (51) and captive dwarf African clawed frogs (Hymenochirus curtipes) (52). As the latter species was widely introduced in ornamental garden ponds throughout the United States in the late 1980s, it may be involved in the dissemination of Batrachochytrium.
Figure 3

Figure 3. Diagrammatic representation of the range of disease outcomes in populations of amphibians affected by a Batrachochytrium-like pathogen. Factors that hypothetically predispose some amphibian populations to declines are illustrated. In this model,...
The ability of a pathogen to cause local population declines resulting in local host extinction requires a mechanism of persistence at low host densities. In epidemiologic models, highly virulent parasites rapidly suppress the host population density below a threshold value required to maintain transmission, resulting in the pathogen's extinction and recovery of the host population (7,10). Microparasites such as Batrachochytrium, with their relatively short duration of infection and high death rates, have an increased threshold population density and are usually less able to persist. Many parasites have evolved life history strategies for persistence (10) and the presence of reservoir hosts may augment the impact of other introduced wildlife diseases on host populations (46). The aclinical presence of Batrachochytrium in the keratinized mouthparts of amphibian larvae implicates this life-cycle stage as a reservoir host for the pathogen. This form of infection may enable Batrachochytrium to persist in reduced amphibian populations (Figure 3). In both Australia and Central America, chytrid infection was observed in larval mouthparts months after initial adult deaths: the larvae of many tropical amphibian species survive 12 to 18 monthsand some temperate species as long as 3 yearsbefore metamorphosing. Examples of larval infection enhancing pathogen-mediated population declines and leading to host population extinctions have been reported for invertebrates (53).
Persistence may be further enhanced by saprophytic development (Figure 3). Batrachochytrium can be cultured in vitro on tryptone agar without the addition of keratin or its derivatives (37,42), and it will grow for at least one generation on cleaned epidermal keratin or on amphibians that have died of the infection (42). Batrachochytrium may survive and reproduce as a saprophytic organism in the environment, at least for short periods. Keratin (from decaying carcasses, shed skin, and other sources) is widely distributed in the environment, and chytrids that use this substrate are well known (42). Furthermore, the ability to develop and reproduce saprophytically is common to many other fungal (54) (including chytrid [41]) and bacterial (55) pathogens. An epidemiologic model of a host-parasite system for pathogens that can reproduce saprophytically clearly shows a lowering of the host threshold population, allowing the pathogen to drive the host to extinction (55). Development of Batrachochytrium for even short periods outside its amphibian host may greatly increase its impact and accelerate population declines. Long-term presence as a saprophyte may explain the lack of recolonization of streams from which amphibians have been extirpated in both Australia (29) and Central America (24,25).
The impact of chytridiomycosis may be enhanced by the ecologic characteristics of certain host species. In Australia, chytridiomycosis-linked deaths have occurred in both declining and nondeclining species (28,29). Species with declining populations belong to a similar ecologic guild: regionally endemic rain forest specialists with low fecundity that reproduce in streams and live at high altitudes (22). These characteristics, which are largely shared by declining Central American amphibians (24,25), are predictors of increased impact from chytridiomycosis. Species that reproduce in streams are probably more susceptible to a waterborne pathogen than terrestrial breeders. Low fecundity (56) and habitat specialization indicate a reduced ability to recover from population declines caused by stochastic events, including disease introduction. Laurance, McDonald, and Speare (39) suggested that the relation between high-altitude populations and declines may be due to a pathogen with a lower preferred developmental temperature. Preliminary data on cultured Batrachochytrium are consistent with this hypothesis: it develops most rapidly at 23oC in culture, with slower growth at 28oC and reversible cessation of growth at 29oC (42). The growth rate of Batrachochytrium in the skin (and therefore virulence) and the survival of zoospores outside the host (and therefore transmission rate) are likely to be lower in amphibians from the warmer lowland regions. The ability of the pathogen to survive saprophytically in the environment and for the disease to persist may also be enhanced in the cooler montane regions. These laboratory data may explain why chytridiomycosis has been associated with population declines in North American amphibians in montane localities (31,32) and after periods of cool weather at many U.S. and Australian sites (29,31).
Multiple factors (host, pathogen, environmental) may be involved in chytridiomycosis emergence. Some authors have hypothesized that infectious disease is only the proximate cause of declines and that environmental factors such as increased UV-B, chemical pollution, climate change, or stress may have predisposed amphibian populations to opportunistic pathogens (13,32,57). Recent work at Monteverde, Costa Rica, suggests that atmospheric warming, with a resultant elevation of the average altitude of the base of the orographic cloud bank and an increase in dry periods, is causally linked to amphibian declines at this site (57). Although no pathologic studies of amphibians were undertaken, overcrowding during periods of drought may have allowed chytridiomycosis to cause substantial deaths (57). Further work is required to test this hypothesis, since a drier climate would also predict a lower overall impact from chytridiomycosisa disease transmitted by flagellated, waterborne zoospores. The evidence suggests that cofactors are not required for chytridiomycosis to cause amphibian mass deaths. Chytridiomycosis is highly pathogenic to captive-bred amphibians exposed in captivity where control animals remained healthy (28,42). Further experimental infections using extremely small inocula (100 zoospores) also proved fatal (29).
Some deaths among wild amphibians have been attributed to immunosuppression, predisposing them to infectious disease (13,32). In the chytridiomycosis-related deaths, chytridiomycosis was consistently found as the cause of death, and the range of opportunistic infections expected to occur in immunocompromised animals was not found (28,29). An increase in UV-B irradiation may influence amphibian declines (58), but in the subtropical regions of Australia and Central America, data demonstrate no significant increase in UV irradiation (29,59). Even so, the potentional effect of increased irradiation on montane riparian rain forest amphibians is uncertain, since these animals lay eggs under rocks or in sand banks and adults are rarely exposed to direct sunlight (24,29). Furthermore, in these regions, the species most likely to be affected by UV increases (arboreal amphibians, which bask or lay exposed eggs) are not in decline (20). Despite extensive research, chemical pollution (20,25), habitat destruction (22), or climate change (57) have not, so far, been causally linked either to the Australian declines or to those at Central American sites other than Monteverde. No other possible cofactors, such as sympatric pathogens, have been found.
Figure 4
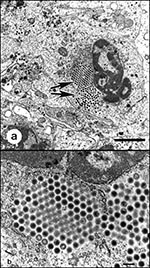
Figure 4. Transmission electron micrographs of iridovirus cultured from the liver of a naturally diseased common frog (Rana temporaria) by using a fathead minnow epithelial cell line. 4a. Virus-infected cell. Large isocahedral viruses...
Iridoviruses have been implicated as the cause of amphibian mass deaths worldwide, with novel iridoviruses of amphibians recently identified from a number of regions (Tables 1, 2). The Iridoviridae encompass five recognized genera: Iridovirus, Chloriridovirus, Ranavirus, Lymphocystivirus, and goldfish virus 1-like viruses (71). Of these, the genus Ranavirus contains pathogens of fish, amphibians, and reptiles (Table 2; Figure 4).
Ranaviruses are often highly virulent and cause systemic infections in amphibians. Experiments with Bohle iridovirus and Gutapo virus suggest that tadpoles are the most susceptible developmental stage for ranavirus infection, and death rates of 100% occur (72). Infected metamorphs die without overt signs of infection, and infected adults show either no overt signs or, occasionally, a general weakness. Histologically, acute necrosis of hematopoietic and lymphoid tissues and of leukocytes occurs in most organs of infected animals (72). Epizootiologic data on tadpole edema virus infections of North American amphibians are scanty, although the virus was isolated mostly from diseased animals (60-62). Experimental infections show that bullfrog (R. catesbeiana) tadpoles infected with tadpole edema virus develop an acute lethal disease after a short incubation. The virus invades the liver, kidneys, and digestive tract and causes hemorrhage in skeletal tissue, pathologic findings similar to those described in B. marinus infected with the ranaviruses frog virus 3, Bohle iridovirus, or Gutapo virus (72). Tadpole edema virus was uniformly lethal to three species of experimentally infected North American toad (61,62). Experimental inoculation with tadpole edema virus caused high rates of deaths in toads (100%, 4 to 17 days postinfection in young and adult Bufo sp. and newly metamorphosed Scaphiopus sp.), 40% death rates in metamorphosing bullfrogs (R. catesbeiana), and no deaths in newly hatched salamanders (Ambystoma sp.). For the last two species, the low virulence suggests either some prior exposure and acquired immunity or a degree of innate resistance.
Pathologic data from ranaviral infections in wild amphibians may be complicated by secondary bacterial infection. Cunningham et al. (26) described two syndromes in Rana temporaria collected at sites of mass deaths in the United Kingdom: a hemorrhagic syndrome affecting the skeletal musculature and the alimentary and reproductive tracts and an ulcerative skin syndrome with dermal ulceration and often necrosis of the distal limbs, but without hemorrhage in the muscles or viscera. In some areas, frogs were found with lesions common to both syndromes (26). These lesions are characteristic of red-leg, a syndrome thought to be caused by bacterial infection; however, Cunningham et al. (26) concluded that the lesions they described were caused by primary iridoviral infection, with or without secondary bacterial infection. Secondary bacterial infection was also reported in wild-collected, diseased A. t. stebbinsi infected with Ambystoma tigrinum virus (27). This infection resulted in rapid onset (5 to 7 days) of apparent epidermal hyperplasia, then dermal hemorrhage, followed by visceral hemorrhage and death rates of up to 45% (27).
The epizootiology of ranaviral disease in amphibians is poorly understood. Data from closely related ranaviral infections of fish, however, suggest a number of factors which may explain their recent emergence. Epizootic hematopoietic necrosis virus is a ranaviral pathogen of fish and the causative agent of epizootic hematopoietic necrosis, a notifiable disease under the Office Internationale des Epizooties (73,74). Sequencing data suggest that this disease is unique among the ranaviruses, is probably indigenous to Australia, and has a wide geographic range. Dissemination may be partly due to the virus's ability to remain infectious under adverse conditions and for prolonged periods (Table 3).
The resistant nature of epizootic hematopoietic necrosis virus suggests that amphibian iridoviruses may survive long periods at the bottom of ponds, particularly during winter. Jancovich et al. (27) demonstrated that water containing Ambystoma tigrinum virus-infected Ambystoma became uninfective after 2 weeks at 25oC, and and epizootics of Ambystoma tigrinum virus and regina ranavirus often followed periods of cold weather (27,36). Epizootic hematopoietic necrosis virus may be spread by fomites such as fishing nets, boats, and fishing rods and through artificial stocking of ponds for recreational fishing (76). Birds have the potential to mechanically transfer virus on their feathers, feet, or bills, or by regurgitation of ingested infected material (76). Similar modes of spread are likely to occur with the amphibian ranaviruses. The occurrence of epizootic hematopoietic necrosis may be correlated with poor husbandry, including overcrowding, inadequate water flow, and fouling of local environments with feed from fish farms (77).
Recent movements of amphibians may have disseminated ranaviral diseases. The North American bullfrog (Rana catesbeiana), the host of tadpole edema virus, has been widely introduced in the western United States and South America. In the United Kingdom, ranaviruses may be disseminated by the common practice of translocation of amphibian egg masses and larvae by humans. Zupanovic et al. (65,78) reported the existence of ranaviruses and antibodies to ranaviruses in the cane toad (Bufo marinus) in Venezuela. In Australia, where B. marinus was introduced to Queensland in 1935, ranaviral antibodies can be identified in this species throughout its range (78). The range of B. marinus has expanded rapidly in Australia, and the toad threatens environmentally sensitive wetlands in the Kakadu National Park. The effects of toxicity, predation, and competition of this relentlessly expanding species are well documented, and its potential to disseminate novel viral pathogens to native amphibians should also be considered in evaluating its ecologic impact. So far, a ranavirus (Bohle iridovirus) has been isolated from only one native Australian amphibian, the ornate burrowing frog (Limnodynastes ornatus) (79).
Experimental infection with Bohle iridovirus causes illness and death in a range of frogs and toads, fish, and reptiles (L. Owens, pers. comm.) (80,81). Gutapo virus, isolated from Venezuelan B. marinus (65), is also able to infect other amphibian species (72). Despite Bohle iridovirus' low host specificity, current levels of surveillance have not implicated it in diseases of free-ranging Australian animals other than those from which it was originally isolated. The ability of Bohle iridovirus to infect fish implicates the widespread introduction of fish for recreational purposes as a potentially significant factor in the dissemination of amphibian ranaviruses. Similarly, Redwood Creek virus, a ranavirus found to cause death of the endangered red-legged frog (67), can infect both amphibians and fish.
The link between amphibian population declines and ranaviral disease is less clear than that with chytridiomycosis. However, these highly virulent ranaviruses are a potential threat to amphibian populations, especially populations isolated from previous disease outbreaks (and thus lacking specific immunity) and species with low fecundity. In particular, the high death rates of tadpoles infected by ranaviruses predict a negative impact on populations. Using the model of Lampo and De Leo (82), a tadpole death rate of 80% to 90% would remove 80% to 90% of the animals that would otherwise survive metamorphosis, resulting in an approximately 80% reduction in adult populations (Lampo, pers. comm.). Although such an epizootic might not endanger the long-term survival of a highly adaptable, highly fecund species such as B. marinus, species that inhabit specialized ecologic niches or have low fecundity might be adversely affected.
Recent outbreaks of iridoviral disease have been particularly notable in Rana temporaria (United Kingdom) and Ambystoma tigrinum (Arizona, Saskatchewan). These outbreaks involved of extensive deaths and the annual recurrence of the diseases. Common ecologic themes occur for these amphibian populations. The natural habitat of A. t. stebbinsi has been entirely destroyed by drainage after human colonization of the southwestern United States, and relict populations are now confined to manmade watering holes for cattle. Other outbreaks of ranaviral disease in A. tigrinum have been associated with altered habitats and artificial ponds. Similar habitat destruction has occurred in the United Kingdom, where most outbreaks of iridovirus infection are reported from artificial ponds (16). High population densities of frogs (16) and salamanders (27) occur in these bodies of water, facilitating the transmission of viruses and predisposing the amphibian populations to mass deaths.
Implications of Emerging Infectious Diseases for Amphibian Population Declines and Wildlife Conservation
The geographic spread of chytridiomycosis threatens populations of endemic and endangered amphibians in rain forests of Australia and Central and South America. Chytridiomycosis has now been reported from 38 amphibian species in 12 families, including ranid and hylid frogs, bufonid toads, and plethodontid salamanders (28,32,37,38,42,83). Although many of these records are from captive animals, the data demonstrate that chytridiomycosis is fatal to species originating from Europe, Africa, Madagascar, the Americas, and Oceania and attest to its potential impact should the disease be introduced into these areas. The emergence of amphibian ranaviruses raises similar concerns. Their ability to infect a wide range of amphibian and fish hosts (different vertebrate classes), global distribution, and high virulence clearly establish them as a global threat to amphibian populations. Despite the recent geographic spread and impact of chytrids and ranaviruses, these diseases probably do not account for all the reports of declining amphibian populations, and evidence exists for other causal factors, such as habitat loss (12). Although chytridiomycosis causes high death rates in a range of species and habitats, it has caused population declines of amphibians only in certain species confined to montane rain forests, while the emergence of ranaviral disease seems to be associated with disturbed or degraded habitats.
Hypotheses regarding the origins and impact of emerging infectious diseases on amphibians are being tested. A number of Batrachochytrium and iridovirus isolates have been cultured, and analysis of their phylogenetic and pathologic features in natural and experimental infections, as well as their biologic characteristics (e.g., ability to survive in the environment, evolution of virulent strains) has already begun. Basic host and parasite ecologic data, such as prevalence and duration of infection and presence of reservoir hosts, are being collected. Data from these studies may enable the formulation of management plans to limit the diseases' impact. Development of chemotherapeutic regimens and diagnostic tests (enzyme-linked immunosorbent assay, in situ hybridization) may enable rapid progress toward these goals (29). National and international structures for the rapid dissemination of information between scientists, politicians, and the public may be crucial in combating the threat of these globally emergent pathogens. However, large geographic areas (e.g., Africa and much of Asia) have not yet been surveyed for declining amphibian populations or for the occurrence of these pathogens. Raising awareness of this threat should be one of the highest priorities for the immediate future.
Many introduced pathogens have affected human populations (10,47). However, far fewer studies of introduced diseases among wildlife have been published, and usually only those producing obvious population losses are studied vigorously (1-6).
The loss of amphibian populations to the point of local extinction is a striking effect of chytridiomycosis in Australia and Central America. However, the effects of introduced wildlife diseases may be more far-reaching and subtle, with knock-on (ripple) effects permeating throughout the ecosystem (1,2,7,82,84). In many tropical and some temperate areas, amphibians make up a significant proportionsometimes mostof these vertebrate biomass (85), and their loss is likely to have unpredictable effects on populations of other species. For example, loss of herbivorous amphibian larvae may cause an overgrowth of algae in montane tropical streams, with further knock-on effects. In Australia and Central America, certain species of snakes prey exclusively on amphibians and are likely to suffer significant population declines, in the same way that the red fox population was drastically reduced in the United Kingdom after the introduction of myxomatosis (10). Such insidious effects following disease introduction and an underestimation of their historical incidence suggest that this pathogen pollution may be as serious a conservation threat as habitat destruction and chemical pollution (1).
Dr. Daszak is a parasitologist at the Institute of Ecology, University of Georgia, Georgia, USA, investigating amphibian chytridiomycosis and other emerging pathogens of wildlife. He is interested in the pathology, cell biology, and host-parasite ecology of emerging diseases of wildlife.
Acknowledgment
The authors thank Trent K. Bollinger, Cynthia Carey, V. Greg Chinchar, Doug Docherty, Timothy Y. James, M. Lampo, Karen Lips, Joyce E. Longcore, Don K. Nichols, David Porter, L. Owens, and Allan P. Pessier for access to unpublished data and papers in press. and Mark A. Farmer, and David Porter, University of Georgia and members of the Infectious Diseases Pathology Activity, Centers for Disease Control and Prevention, for their hospitality.
References
- Daszak P, Cunningham AA, Hyatt AD. Emerging infectious diseases of wildlife—threats to human health and biodiversity. Science. 1999. In press.
- Lyles AM, Dobson AP. Infectious disease and intensive management: population dynamics, threatened hosts, and their parasites. J Zoo Wildl Med. 1993;24:315–26.
- Viggers KL, Lindenmayer DB, Spratt DM. The importance of disease in reintroduction programmes. Wildl Res. 1993;20:687–98. DOIGoogle Scholar
- Cunningham AA. Disease risks of wildlife translocations. Conserv Biol. 1996;10:349–53. DOIGoogle Scholar
- Plowright W. The effects of rinderpest and rinderpest control on wildlife in Africa. Symposia of the Zoological Society of London. 1982;50:1–28.
- Harvell CD, Kim K, Bukholder JM, Colwell RR, Epstein PR, Grimes DJ, Emerging marine diseases—climate links and anthropogenic factors. Science. 1999;285:1505–10. DOIPubMedGoogle Scholar
- Dobson AP, May RM. Disease and conservation. In: Soulé M, editor. Conservation biology: The science of scarcity and diversity. Sunderland (MA): Sinauer Assoc Inc.; 1986. p. 345-65.
- Scott ME. The impact of infection and disease on animal populations: Implications for conservation biology. Conserv Biol. 1988;2:40–56. DOIGoogle Scholar
- Daszak P, Cunningham AA. Wildlife disease biology comes of age. Oryx. 1998;32:238–9. DOIGoogle Scholar
- Anderson RM, May RM. The invasion, persistence and spread of infectious diseases within animal and plant communities. Philos Trans R Soc Lond B Biol Sci. 1986;314:533–70. DOIPubMedGoogle Scholar
- Tompkins DM, Begon M. Parasites can regulate wildlife populations. Parasitol Today. 1999;15:311–3. DOIPubMedGoogle Scholar
- Blaustein AR, Wake DB. Declining amphibian populations: A global phenomenon. Trends Ecol Evol. 1990;5:203–4. DOIGoogle Scholar
- Carey C. Hypothesis concerning the causes of the disappearance of boreal toads from the mountains of Colorado. Conserv Biol. 1993;7:355–62. DOIGoogle Scholar
- Drost CA, Fellers GM. Collapse of a regional frog fauna in the Yosemite area of the California Sierra Nevada, USA. Conserv Biol. 1996;10:414–25. DOIGoogle Scholar
- Cunningham AA, Langton TES, Bennett PM, Drury SEN, Gough RE, Kirkwood JK. Unusual mortality associated with poxvirus-like particles in frogs (Rana temporaria). Vet Rec. 1993;133:141–2.PubMedGoogle Scholar
- Cunningham AA, Langton TES, Bennett PM, Lewin JF, Drury SEN, Gough RE, Investigations into unusual mortalities of the common frog (Rana temporaria) in Britain. In: Proceedings of the 5th International Colloquium on the Pathology of Reptiles and Amphibians; 1995 March 3; The Netherlands. p. 19-27.
- Gupta BK. Declining amphibians. Curr Sci. 1998;75:81–4.
- Weygoldt P. Changes in the composition of mountain stream frog communities in the Atlantic Mountains of Brazil: Frogs as indicators of environmental deteriorations? Stud Neotrop Fauna Environ. 1989;24:249–55. DOIGoogle Scholar
- Mahony M. The decline of the green and golden bell frog Litoria aurea viewed in the context of declines and disappearances of other Australian frogs. Aust Zool. 1996;30:237–47.
- Richards SJ, McDonald KR, Alford RA. Declines in populations of Australia's endemic tropical forest frogs. Pac Conserv Biol. 1993;1:66–77.
- Williams SE, Hero J-M. Rainforest frogs of the Australian Wet Tropics: Guild classification and the ecological similarity of declining species. Proc R Soc Lond B Biol Sci. 1998;265:597–602. DOIGoogle Scholar
- Pounds JA, Fogden MPL, Savage JM, Gorman GC. Tests of null models for amphibian declines on a tropical mountain. Conserv Biol. 1997;11:1307–22. DOIGoogle Scholar
- Lips KR. Mass mortality and population declines of anurans at an upland site in Western Panama. Conserv Biol. 1999;13:117–25. DOIGoogle Scholar
- Cunningham AA, Langton TES, Bennett PM, Lewin JF, Drury SEN, Gough RE, Pathological and microbiological findings from incidents of unusual mortality of the common frog (Rana temporaria). Philos Trans R Soc Lond B Biol Sci. 1996;351:1529–57. DOIPubMedGoogle Scholar
- Jancovich JK, Davidson EW, Morado JF, Jacobs BL, Collins JP. Isolation of a lethal virus from the endangered tiger salamander Ambystoma tigrinum stebbinsi. Dis Aquat Organ. 1997;31:161–7. DOIGoogle Scholar
- Berger L, Speare R, Daszak P, Green DE, Cunningham AA, Goggin CL, Chytridiomycosis causes amphibian mortality associated with population declines in the rainforests of Australia and Central America. Proc Natl Acad Sci U S A. 1998;95:9031–6. DOIPubMedGoogle Scholar
- Berger L, Speare R, Hyatt AD. Chytrid fungi and amphibian declines: Overview, implications and future directions. In: Campbell A, editor. Declines and disapparances of Australian frogs. Environmental Australia, Canberra, Australia: Environmental Australia; 2000. p.21-31.
- Daszak P, Cunningham AA. Extinction by infection. Trends Ecol Evol. 1999;14:279. DOIPubMedGoogle Scholar
- Carey C, Cohen N, Rollins-Smith L. Amphibian declines: an immunological perspective. Dev Comp Immunol. 1999;23:459–72. DOIPubMedGoogle Scholar
- Drury SEN, Gough RE, Cunningham AA. Isolation of an iridovirus-like agent from common frogs (Rana temporaria). Vet Rec. 1995;137:72–3.PubMedGoogle Scholar
- Hyatt AD, Gould AR, Zupanovic Z, Cunningham AA, Hengstberger S, Whittington RJ, Characterisation of piscine and amphibian iridoviruses. Arch Virol. 2000. In press.PubMedGoogle Scholar
- Docherty D, Chinchar VG, Meteyer C, Brannian R, Hansen W, Wang J, A diagnostic evaluation of three salamander mortality events associated with an iridovirus and subsequent genomic comparison of the virus isolates obtained. In: Proceedings of the 48th Annual Wildlife Disease Association Conference; 1999 Aug 8-12; Athens, Georgia.
- Bollinger TK, Mao J, Schock D, Brigham RM, Chinchar VG. Pathology, isolation and preliminary molecular characterization of a novel iridovirus from tiger salamanders in Saskatchewan. J Wildl Dis. 1999;35:413–29.PubMedGoogle Scholar
- Pessier AP, Nichols DK, Longcore JE, Fuller MS. Cutaneous chytridiomycosis in poison dart frogs (Dendrobates spp.) and White's tree frogs (Litoria caerulea). J Vet Diagn Invest. 1999;11:194–9.PubMedGoogle Scholar
- Milius S. Fatal skin fungus found in U.S. frogs. Science News 1998;July 4th:7.
- Laurance WF, McDonald KR, Speare R. Epidemic disease and the catastrophic decline of Australian rain forest frogs. Conserv Biol. 1996;10:406–13. DOIGoogle Scholar
- Alford RA, Richards SJ. Lack of evidence for epidemic disease as an agent in the catastrophic decline of Australian rain forest frogs. Conserv Biol. 1997;11:1026–9. DOIGoogle Scholar
- Powell MJ. Looking at mycology with a Janus face: A glimpse at chytridiomycetes active in the environment. Mycologia. 1993;85:1–20. DOIGoogle Scholar
- Longcore JE, Pessier AP, Nichols DK. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia. 1999;91:219–27. DOIGoogle Scholar
- Garnett GP, Homes EC. The ecology of emergent infectious disease. Bioscience. 1996;46:127–35. DOIGoogle Scholar
- Laurance WF, McDonald KR, Speare R. In defense of the epidemic disease hypothesis. Conserv Biol. 1997;11:1030–4. DOIGoogle Scholar
- McCallum H, Dobson A. Detecting disease and parasite threats to endangered species and ecosystems. Trends Ecol Evol. 1995;10:190–4. DOIGoogle Scholar
- Dobson AP, Carper ER. Infectious diseases and human population history. Bioscience. 1996;46:115–26. DOIGoogle Scholar
- Burdon JJ. Fungal pathogens as selective forces in plant populations and communities. Aust J Ecol. 1991;16:423–32. DOIGoogle Scholar
- Vitousek PM, D'Antonio CM, Loope LL, Westbrooks R. Biological invasions as global environmental change. Am Sci. 1996;84:468–78.
- Scott NJ. Postmetamorphic death syndrome. Froglog. 1993;7:1–2.
- Taylor SK, Williams ES, Thorne ET, Mills KW, Withers DI, Pier AC. Causes of mortality of the Wyoming toad. J Wildl Dis. 1999;35:49–57.PubMedGoogle Scholar
- Groff JM, Mughannam A, McDowell TS, Wong A, Dykstra MJ, Frye FL, An epizootic of cutaneous zygomycosis in cultured dwarf African clawed frogs (Hymenochirus curtipes) due to Basidiobolus ranarum. J Med Vet Mycol. 1991;29:215–23. DOIPubMedGoogle Scholar
- Richard A, Cory J, Speight M, Williams T. Foraging in a pathogen reservoir can lead to local host population extinction: A case study of lepidoptera-virus interaction. Oecologia. 1999;118:29–38. DOIPubMedGoogle Scholar
- Cox RA. Coccidioidomycosis. In: Fungal infections and immune responses. Murphy JW, Friedman H, Bendinelli M, editors. New York: Plenum Press; 1993. p. 173-211.
- Godfray HCJ, Briggs CJ, Barlow ND, O'Callaghan M, Glare TR, Jackson TA. A model of insect-pathogen dynamics in which a pathogenic bacterium can also reproduce saprophytically. Proc R Soc Lond B Biol Sci. 1999;266:233–40. DOIGoogle Scholar
- Bennett PM, Owens IPF. Variation in extinction risk among birds: Chance or evolutionary predisposition? Proc R Soc Lond B Biol Sci. 1997;264:401–8. DOIGoogle Scholar
- Pounds JA, Fogden MPL, Campbell JH. Biological response to climate change on a tropical mountain. Nature. 1999;398:611–5. DOIGoogle Scholar
- Kiesecker JM, Blaustein AR. Synergism between UV-B radiation and a pathogen magnifies amphibian embryo mortality in nature. Proc Natl Acad Sci U S A. 1995;92:11049–52. DOIPubMedGoogle Scholar
- Madronich S, de Gruijl FR. Skin cancer and UV radiation. Nature. 1993;366:23. DOIPubMedGoogle Scholar
- Granoff A, Came PE, Rafferty KA. The isolation and properties of viruses from Rana pipiens: Their possible relationship to the renal adenocarcinoma of the leopard frog. Ann N Y Acad Sci. 1965;126:237–55. DOIPubMedGoogle Scholar
- Clark HF, Brennan JC, Zeigel RF. Karzon. Isolation and characterization of viruses from the kidneys of Rana pipiens with renal adenocarcinoma before and after passage in red eft (Triturus viridescens). J Virol. 1968;2:629–40.PubMedGoogle Scholar
- Wolf K, Bullock GL, Dunbar CE, Quimby MC. Tadpole edema virus: A viscerotrophic pathogen for anuran amphibians. J Infect Dis. 1968;118:253–62.PubMedGoogle Scholar
- Fijan N, Matasin Z, Petrinec Z, Valpotic I, Zwillenberg LO. Isolation of an iridovirus-like agent from the green frog Rana esculenta L. Veterinarski Archiv Zagreb. 1991;61:151–8.
- Hengstberger SG, Hyatt AD, Speare R, Coupar BEH. Comparison of epizootic haematopoietic necrosis and Bohle iridoviruses, recently isolated Australian iridoviruses. Dis Aquat Organ. 1993;15:93–107. DOIGoogle Scholar
- Zupanovic Z, Musso C, Lopez G, Louriero C-L, Hyatt AD, Hengstberger S, Isolation and characterization of iridoviruses from the giant toad Bufo marinus in Venezuela. Dis Aquat Organ. 1998;33:1–9. DOIPubMedGoogle Scholar
- Mao J, Green DE, Fellers G, Chinchar VG. Molecular characterization of iridoviruses isolated from sympatric amphibians and fish. Virus Res. 1999;63:45–52. DOIPubMedGoogle Scholar
- Mao J, Hedrick RP, Chinchar VG. Molecular characterization, sequence analysis and taxonomic position of newly isolated fish iridoviruses. Virology. 1997;229:212–20. DOIPubMedGoogle Scholar
- Zhang QY, Li ZQ, Gui JF. Studies on morphogenesis and cellular interactions of Rana grylio virus in an infected fish cell line. Aquaculture. 1999;175:185–97. DOIGoogle Scholar
- Westhouse RA, Jacobsen ER, Harris RK, Winter KR, Homer BL. Respiratory and pharyngo-esophageal iridovirus infection in a gopher tortoise (Gopherus polyphemus). J Wildl Dis. 1996;32:682–6.PubMedGoogle Scholar
- Chen Z-X, Zheng J-C, Jiang Y-I. A new iridovirus from soft-shelled turtle. Virus Res. 1999;63:147–51. DOIPubMedGoogle Scholar
- Murphy FA, Fauquet CM, Bishop DHL, Ghabrial SA, Jarvis AW, Martelli P, Iridoviridae. In: Murphy FA, Fauquet CM, Bishop DHL, Ghabrial SA, Jarvis AW, Martelli P, et al., editors. Virus taxonomy: The classification and nomenclature of viruses. The sixth report of the international committee on the taxonomy of viruses. New York, Wien: Springer-Verlag; 1995. p. 95-9.
- Hyatt AD, Parkes H, Zupanovic Z. Identification, characterisation and assessment of Venezuelan viruses for potential use as biological control agents against the cane toad (Bufo marinus) in Australia. Report to the Australian Federal Government and Environment Australia. 1998.
- Ahne W, Bremont M, Hedrick RP, Hyatt AD, Whittington RJ. Iridoviruses associated with epizootic haematopoietic necrosis (EHN) in aquaculture. World J Microbiol Biotechnol. 1997;13:367–73. DOIGoogle Scholar
- Hyatt AD, Whittington RJ. National diagnostic tests for the detection of epizootic haematopoietic necrosis virus (EHNV) and certification of EHNV-free fish. Final report (project 92/66). Fish Research Development Council of Australia; 1997.
- Langdon JS. Experimental transmission and pathogenicity of epizootic haematopoietic necrosis virus (EHNV) in redfin perch, Perca fluviatilis L., and 11 other teleosts. J Fish Dis. 1989;12:295–310. DOIGoogle Scholar
- Whittington RJ, Kearns C, Hyatt AD, Hengstberger S, Rutzou T. Spread of epizootic haematopoietic necrosis virus (EHNV) in redfin perch (Perca fluviatilis) in Southern Australia. Aust Vet J. 1996;73:112–4. DOIPubMedGoogle Scholar
- Whittington RJ, Hyatt AD. Diagnosis and prevention of epizootic haematopoietic necrosis virus infection. Proceedings of the National Research Institute of Aquaculture International Workshop: New approaches to viral diseases of aquatic animals; 1997 Jan 21-24; Kyoto, Japan. National Research Institute of Aquaculture; 1997.
- Zupanovic Z, Lopez B, Hyatt AD, Green B, Bartran G, Parkes H, Giant toads Bufo marinus in Australia and Venezuela have antibodies against "ranaviruses". Dis Aquat Organ. 1998;32:1–8. DOIPubMedGoogle Scholar
- Speare R, Smith JR. An iridovirus-like agent isolated from the ornate burrowing frog Limnodynastes ornatus in northern Australia. Dis Aquat Organ. 1992;14:51–7. DOIGoogle Scholar
- Cullen BR, Owens L, Whittington RJ. Experimental infection of Australian anurans (Limnodynastes terraereginae and Litoria latopalmata) with Bohle iridovirus. Dis Aquat Organ. 1995;23:83–92. DOIGoogle Scholar
- Moody NJC, Owens L. Experimental demonstration of the pathogenicity of a frog virus, Bohle iridovirus, for a fish species, barramundi Lates calcarifer. Dis Aquat Organ. 1994;18:95–102. DOIGoogle Scholar
- Lampo M, De Leo GA. The invasion ecology of the toad Bufo marinus: From South America to Australia. Ecol Appl. 1998;8:388–96.
- Nichols DK, Pessier AP, Longcore JE. Cutaneous chytridiomycosis: an emerging disease? Proceedings American Association of Zoo Veterinarians 1998:269-71.
- Lessios HA. Mass mortality of Diadema antillarum in the Caribbean: What have we learned? Annu Rev Ecol Syst. 1988;19:371–93.
- Burton TM, Likens GE. Salamander populations and biomass in the Hubbard Brook Experimental Forest, New Hampshire. Copeia. 1975;3:541–6. DOIGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 5, Number 6—December 1999
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Peter Daszak, Institute of Ecology, University of Georgia, Ecology Building, Athens, GA 30602, USA; fax: 706-542-4819
Top