Volume 12, Number 2—February 2006
Dispatch
Introductions of West Nile Virus Strains to Mexico
Cite This Article
Citation for Media
Abstract
Complete genome sequencing of 22 West Nile virus isolates suggested 2 independent introductions into Mexico. A previously identified mouse-attenuated glycosylation variant was introduced into southern Mexico through the southeastern United States, while a common US genotype appears to have been introduced incrementally into northern Mexico through the southwestern United States.
West Nile virus (WNV), a mosquitoborne flavivirus for which birds serve as reservoir and amplification hosts, was introduced into New York in 1999 (1) and spread across the United States to California by 2003 (2). By 2002, serosurveys demonstrated WNV circulation in >6 eastern Mexican states and along its northern border with the United States (3–5). This pattern of WNV appearance in Mexico suggested a southwesterly spread across the United States and into northeastern Mexico through Texas. However, in the spring of 2003, the first WNV isolate found in Mexico was obtained from a raven in the southeastern state of Tabasco (3). If WNV reached southern Mexico by incremental spread through northern Mexico from Texas, the index isolate would have been expected sooner and in a northern Mexican state. Phylogenetic analyses showed the raven isolate to be unexpectedly divergent from contemporary Texas strains, but exact relationships and a route of entry could not be determined by using premembrane and envelope glycoprotein (prM-E) sequences (3).
The divergence between the southern Mexican raven and Texas isolates suggested that WNV arrived in southern Mexico by an alternate route, perhaps the Caribbean. After its spread throughout the northeastern United States, WNV appeared abruptly in Florida in 2001, appearing to bypass several mid-Atlantic states. This pattern could be explained by spread of migratory birds (6); the Atlantic coast flyway overlaps both New York and Florida, while the Mississippi Valley flyway overlaps both Louisiana and the Yucatan Peninsula of Mexico. Longitudinal avian serosurveys that began in 2000 showed WNV seropositivity in at least 3 migratory and 2 resident bird species captured in the Yucatan Peninsula from 2002 to 2003 (7). Thus, migratory birds may have carried WNV from the southeastern United States into Mexico, either directly or through the Caribbean. Serosurveys have suggested WNV circulation among birds in various Caribbean islands since 2002 (8–10).
The possibility of a third WNV introduction into Mexico at the California border must also be considered. A 2003 horse isolate from the northern Mexican state of Nuevo Leon was closely related to Texas isolates from 2002 (11), based on its prM-E sequence. We do not know whether WNV reached California from Texas and the Midwest by crossing the Rocky Mountains or by traveling first into northern Mexico and subsequently spreading north from Baja California. The latter route is suggested by the geographic link with the first detection of WNV in southeastern California (2).
The reported incidence of human West Nile encephalitis is much greater on the US (California) than on the Mexican (Baja California and Sonora) side of the common border. Possible explanations for this discrepancy include differences in disease surveillance and reporting. Another possibility is that the WNV strains circulating in Mexico are attenuated compared to US strains, and the identification of a murine-attenuated glycosylation variant in Tabasco State (12) is consistent with this hypothesis.
Figure 1
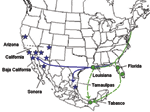
To investigate possible routes of WNV entry into Mexico from the United States, 9 isolates from Mexico (all strains available) and 13 strains isolated in the United States from hypothetical points of introduction into Mexico (2 from Florida, 2 from Louisiana, 3 from Arizona, and 6 from California) were compared. Isolates from several northern Mexican states, 1 from Sonora, 1 from Tamaulipas, and 7 from Baja California, were obtained from a variety of birds and from a horse (Table, Figure 1) by injection of Vero cells. RNA was extracted from first or second Vero cell passages by using the QIAamp Viral RNA Mini-kit (Qiagen Inc, Valencia, CA, USA). Reverse transcription–polymerase chain reactions (RT-PCRs) were performed to amplify the complete WNV genome in 6 overlapping amplicons by using primers described previously (12). Amplicons were purified from agarose gels by using the QIAquick gel-extraction kit (Qiagen), and both strands were sequenced directly by using the PCR primers and the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied BioSystems, Foster City, CA, USA) with a 3100 Genetic Analyzer (Applied Biosystems).
Complete genomic sequences excluding the 5´ and 3´ terminal 20 nucleotides (representing primers incorporated into amplicons) were aligned with all homologous WNV sequences from the GenBank library by using ClustalW. Sequences were analyzed by using maximum parsimony and neighbor-joining programs implemented in the PAUP 4.0 software package (13) as well as Bayesian analysis using MRBAYES v3.0 (14) with 100,000 generations, a general time-reversible model with empirically estimated base frequencies, and either a codon position-specific (for the open reading frame alone) or a gamma distribution of substitution rates among nucleotide sites.
Figure 2
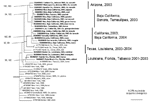
Figure 2. Phylogenetic tree generated from the complete open reading frame of West Nile virus sequences using a Bayesian analysis. Virus strains are labeled by GenBank accession number followed by the state and/or...
All phylogenetic trees placed the North American WNV isolates into monophyletic groups with strong bootstrap and Bayesian support values; the tree generated using the Bayesian analyses is presented in Figure 2. The Tabasco 2003 isolate grouped with 3 strains from 2001 and 2002 in Florida and Louisiana and more distantly with a New York isolate from 2000, with strong Bayesian probability and bootstrap support; inclusion of the New York grouse strain was weakly supported (bootstrap and Bayesian probability values <80%). In contrast, a 2004 Louisiana isolate and other recent strains from Texas were positioned basally to the large clade containing all northern Mexico, California, and Arizona isolates. This California/Arizona/northern Mexico group was highly conserved, with <0.5% nucleotide and 0.04% amino acid sequence divergence. The 2003 Tabasco strain was phylogenetically distinct from all other Mexico isolates, which grouped with the California and Arizona isolates. Surprisingly, despite the greater geographic distances between Tamaulipas and Baja California/Sonora, compared to the distance between Tamaulipas and Texas, the Tamaulipas WNV strains grouped more closely with strains from Baja California and Sonora than with those from Texas.
Compared to the Tabasco strain, the other Mexican isolates differed by 0.55%–0.66% nucleotide sequence divergence across the genome. The gene with the most sequence divergence was prM, with 0.72%–1.4% divergence from the Tabasco strain. However, the 5´ untranslated region was more variable with 3.0%–4.6% divergence. The most conserved gene was NS2B, with 0.0%–0.24% divergence from the Tabasco strain. The E gene, often used for phylogenetic analyses, had 0.46%–0.66% sequence divergence.
Of the Mexican WNV isolates, only the 2003 Tabasco raven isolate had the E-156 Pro residue, which ablates the N-linked glycosylation site found in most North American strains. In addition, 2 other WNV isolates (GenBank accession nos. AY490240 and AF260968) share this E-156 Pro residue despite their geographic diversity (China and Egypt, respectively) and their placement in different lineages. Although the paraphyletic nature of this Pro mutation suggests that it could be selected either during laboratory isolation or passage, its identification in the low-passage Tabasco isolate may indicate its presence in nature.
Our data support the hypothesis that WNV entered Mexico through at least 2 independent introductions. The introduction detected by the first virus isolation in May 2003 from a raven in Tabasco State probably occurred from a migratory bird that flew southward from the southeastern United States to the Gulf of Mexico or the Caribbean and bypassed northern Mexico. The isolation and sequencing of WNV isolates from islands in the Caribbean may shed further light on how WNV reached southern Mexico. However, the extreme genetic conservation of North American WNV strains may preclude identifying the exact routes of introduction. Independently, other WNV strains probably spread incrementally from the southwestern United States into northern Mexico. Both northward and southward movements of WNV between northern Mexico and California or Arizona may also occur.
Available WNV strains from Mexico indicate that the murine-attenuated, E-156 glycosylation-negative variant identified in Tabasco state may be limited in its distribution to southern Mexico, while the glycosylated variant typical of US strains is widespread in northern Mexico. However, our sampling was limited and may also be biased because many WNV isolates were from sick or dying animals; the attenuated E-156 Pro residue phenotype could be undersampled because relatively benign infections are rarely identified.
The epidemiology of WNV-associated disease in Mexico is puzzling. According to the Centers for Disease Control and Prevention, 2,470 human cases of WNV infection were confirmed during 2004 in the United States, with >80% of these from areas of California and Arizona bordering the northern Mexico states of Baja California and Sonora where many of our viral isolations were made. In contrast, only 7 human cases of WNV have been confirmed in Mexico. The cases occurred in the border states of Chihuahua (n = 4), Sonora (n = 1), and Nuevo Leon (n = 1) in 2003, and Sonora (n = 1) in 2004 (15). Our results of extremely low sequence divergence between the southwestern United States and northern Mexican WNV isolates indicate that this epidemiologic discrepancy is unlikely to be explained by genetic and phenotypic differences among WNV strains. The possibility that WNV circulating in Mexico has an attenuated phenotype was suggested by the murine-attenuating mutation in the Tabasco raven isolate (12). However, none of our northern Mexico isolates have the E-156-P attenuating mutation, and all appear extremely closely related to isolates made in southwestern areas of the United States with a high disease incidence.
Another possible explanation for the low incidence of WNV disease in Mexico is resistance in the Mexican human population, possibly because cross-protective immunity from other flavivirus infections such as dengue and St. Louis encephalitis viruses. Although St. Louis encephalitis virus is common in some areas of the continental United States, including California, dengue virus infections are rare; only 157 cases of dengue were reported in the northern states of Mexico in 2004: 25 in Sonora; 21 in Nuevo Leon; 88 in Tamaulipas; 3 in Coahuila; 0 in Chihuahua; and 0 in Baja California (http://www.dgepi.salud.gob.mx/boletin/2004/sem52). Of all Mexican states, Baja California and Sonora adjacent to the US border have the lowest incidence of flaviviral infections. Human flavivirus serosurveys should be conducted in northern Mexico to further evaluate the possibility of cross-protective flavivirus immunity. Newer approaches to detect and identify flaviviral disease are also needed in Mexico to more accurately assess the impact of WNV.
Ms Deardorff is a graduate student in the experimental pathology program at the University of Texas Medical Branch. Her research interests include arbovirology, zoonoses, viral ecology, and evolution.
Acknowledgments
We thank Igor Romero and many veterinarians from CPA-SAGARPA for field support; Emily N. Green for technical assistance; and Ronald C. Cheshier, Lilian Stark, and Barbara Cahoon-Young for providing virus isolates.
This research was supported by contract N01-AI25489 from the National Institutes of Health and by the California Mosquito Research Program. E.D. was supported by the T01/CCT622892 CDC Fellowship Training Grant in Vector-Borne Infectious Diseases. C.T.D. was supported by NIH T32 training grant AI 7256 in Emerging and Tropical Infectious Diseases.
References
- Lanciotti RS, Roehrig JT, Deubel V, Smith J, Parker M, Steele K, Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science. 1999;286:2333–7. DOIPubMedGoogle Scholar
- Reisen W, Lothrop H, Chiles R, Madon M, Cossen C, Woods L, West Nile virus in California. Emerg Infect Dis. 2004;10:1369–78.PubMedGoogle Scholar
- Estrada-Franco JG, Navarro-Lopez R, Beasley DW, Coffey L, Carrara AS, Travassos da Rosa A, West Nile virus in Mexico: evidence of widespread circulation since July 2002. Emerg Infect Dis. 2003;9:1604–7.PubMedGoogle Scholar
- Lorono-Pino MA, Blitvich BJ, Farfan-Ale JA, Puerto FI, Blanco JM, Marlenee NL, Serologic evidence of West Nile virus infection in horses, Yucatan State, Mexico. Emerg Infect Dis. 2003;9:857–9.PubMedGoogle Scholar
- Blitvich BJ, Fernandez-Salas I, Contreras-Cordero JF, Marlenee NL, Gonzalez-Rojas JI, Komar N, Serologic evidence of West Nile virus infection in horses, Coahuila State, Mexico. Emerg Infect Dis. 2003;9:853–6.PubMedGoogle Scholar
- Rappole JH, Hubalek Z. Migratory birds and West Nile virus. J Appl Microbiol. 2003;94(Suppl):47S–58S. DOIPubMedGoogle Scholar
- Farfan-Ale JA, Blitvich BJ, Lorono-Pino MA, Marlenee NL, Rosado-Paredes EP, Garcia-Rejon JE, Longitudinal studies of West Nile virus infection in avians, Yucatan State, Mexico. Vector Borne Zoonotic Dis. 2004;4:3–14. DOIPubMedGoogle Scholar
- Dupuis AP II, Marra PP, Kramer LD. Serologic evidence of West Nile virus transmission, Jamaica, West Indies. Emerg Infect Dis. 2003;9:860–3.PubMedGoogle Scholar
- Komar O, Robbins MB, Klenk K, Blitvich BJ, Marlenee NL, Burkhalter KL, West Nile virus transmission in resident birds, Dominican Republic. Emerg Infect Dis. 2003;9:1299–302.PubMedGoogle Scholar
- Quirin R, Salas M, Zientara S, Zeller H, Labie J, Murri S, West Nile virus, Guadeloupe. Emerg Infect Dis. 2004;10:706–8.PubMedGoogle Scholar
- Blitvich BJ, Fernandez-Salas I, Contreras-Cordero JF, Lorono-Pino MA, Marlenee NL, Diaz FJ, Phylogenetic analysis of West Nile virus, Nuevo Leon State, Mexico. Emerg Infect Dis. 2004;10:1314–7.PubMedGoogle Scholar
- Beasley DW, Davis CT, Estrada-Franco J, Navarro-Lopez R, Campomanes-Cortes A, Tesh RB, Genome sequence and attenuating mutations in West Nile virus isolate from Mexico. Emerg Infect Dis. 2004;10:2221–4.PubMedGoogle Scholar
- Swofford DL. 1998. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4, Sunderland (MA), Sinauer Associates.
- Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001;17:754–5. DOIPubMedGoogle Scholar
- Ramos C, Falcon-Lezama A. La fiebre del Nilo Occidental: una enfermedad emergente en Mexico. Salud Publica Mex. 2004;46:488–90. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 12, Number 2—February 2006
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Scott C. Weaver, Department of Pathology, University of Texas Medical Branch, Galveston, TX 77555-0609, USA; fax: 409-747-2415
Top