Volume 12, Number 8—August 2006
Dispatch
Community-acquired Methicillin-resistant Staphylococcus aureus in Children, Taiwan
Cite This Article
Citation for Media
Abstract
Highly virulent community-acquired methicillin-resistant Staphylococcus aureus (CA-MRSA) with Panton-Valentine leukocidin (PVL) is common worldwide. Using antimicrobial drug susceptibility testing, staphylococcal cassette chromosome mec typing, exotoxin profiling, and pulsed-field gel electrophoresis typing, we provide evidence that supports the relationship between nasal strains of PVL-positive MRSA and community-acquired disease.
Panton-Valentine leukocidin (PVL) is a 2-component cytotoxin that targets human and rabbit polymorphonuclear cells, monocytes, and macrophages (1). Gene products of PVL (lukS-PV and lukF-PV), which are encoded by contiguously located, cotranscribed genes (lukS-PV and lukF-PV), assemble as hetero-oligomers and synergistically exert cytolytic pore-forming activity (1). PVL is mainly associated with primary cutaneous infections, especially furuncles, and with severe necrotizing community-acquired pneumonia (2). The PVL locus is present in most community-acquired methicillin-resistant Staphylococcus aureus (CA-MRSA) isolates studied and is a stable marker of CA-MRSA strains worldwide (3).
In a previous study, we found that CA-MRSA skin and soft tissue infections among Taiwanese children are caused by a predominantly endemic strain that has PVL genes (4). Nasal carriage of MRSA also plays a key role in the epidemiology and pathogenesis of community-associated disease (5). Therefore, our prospective investigation sought to 1) determine the prevalence of PVL-positive S. aureus among isolates from children with various staphylococcal diseases and asymptomatic nasal colonization, and 2) test the hypothesis that CA-MRSA infection is associated with community PVL-positive MRSA nasal carriage.
The study protocol was reviewed and approved by the National Defense Medical Center Institutional Review Board. Two collections of S. aureus isolates were used. A list of all children <14 years of age hospitalized with various staphylococcal infections during the period from December 2003 to November 2005 was compiled from records at the clinical microbiology laboratory at the Tri-Service General Hospital in Taipei. The first collection of 144 infecting strains was further categorized into 11 types of staphylococcal infection on the basis of the clinical details provided. A case was considered community acquired if MRSA was isolated from cultures of specimens obtained within 72 hours after hospitalization. Risk factors for MRSA infection included hospitalization <12 months before the date of MRSA isolation; history of any surgical procedure; history of endotracheal intubation; underlying chronic disorder; antimicrobial drug therapy <12 months before the date of MRSA isolation; presence of an indwelling venous or urinary catheter; or household contact with a person with an identified risk factor or a worker in a healthcare environment (6).
A second collection of 300 colonizing strains was obtained during the same period by culturing samples from the anterior nares of 1,195 healthy children in the community. Eligible participants were <14 years of age with no acute medical problem who either visited a healthcare facility for a well-child checkup or attended 1 of 7 kindergartens in Taipei.
MRSA identification and antimicrobial drug susceptibility were determined according to the Clinical Laboratory Standards Institute (formerly known as the National Committee for Clinical Laboratory Standards) guidelines (7,8). Staphylococcal cassette chromosome mec (SCCmec) elements were typed and PVL genes were detected as described (2,9,10). Sequences specific for sea to see, seg to sei, eta, etb, and tst, which encode staphylococcal enterotoxins (SEA to SEE, and SEG to SEI), exfoliative toxins (ETA and ETB), and toxic shock syndrome toxin-1, respectively, were detected by using methods described by Jarraud et al. (11). Pulsed-field gel electrophoresis (PFGE) was performed by using a CHEF Mapper XA system (Bio-Rad Laboratories, Hercules, CA, USA) according to a published protocol (12).
Data were analyzed by the Mantel-Haenszel test and χ2 test with SPSS version 10.0 software (SPSS, Chicago, IL, USA). A p value <0.05 was considered significant.
Of the 444 isolates examined, PVL-positive isolates constituted 23% of all S. aureus isolates analyzed (Table 1). Among 144 isolates (67 methicillin-susceptible S. aureus isolates and 77 MRSA isolates) from different staphylococcal infections, 82 (56.9%) were PVL positive, and most were associated with skin and soft tissue infections (especially furuncles). In contrast, only 18 (6%) of 300 colonizing isolates had the PVL locus. PVL genes were also found in isolates associated with other deep-space infections including pyomyositis, osteomyelitis, and septic arthritis. Of 1,195 healthy children who were screened, 89 (7.4%) had cultures with MRSA. Only 15 (16.9%) of 89 community MRSA-colonizing strains were PVL positive.
Among the 144 S. aureus isolates obtained from various types of staphylococcal disease, 32 (22.2%) isolates were further confirmed as CA-MRSA according to the inclusion criteria (abscess [n = 5], carbuncles [n = 6], cellulitis [n = 9], furuncles [n = 3], pyomyositis [n = 1], osteomyelitis [n = 1], pneumonia [n = 1], and staphylococcal scarlet fever [n = 6]); all contained the genes encoding PVL. Antibiograms of 15 PVL-positive MRSA-colonizing strains did not differ significantly from those of 32 CA-MRSA–infecting strains with respect to clindamycin, erythromycin, gentamicin, and chloramphenicol. All 47 PVL-positive MRSA isolates were resistant to penicillin G, and none had reduced susceptibility to vancomycin, teicoplanin, trimethoprim-sulfamethoxazole, fusidic acid, or ciprofloxacin.
To gain insight into the association between nasal strains of PVL-positive MRSA and community-acquired disease, we designed a comparative study in which the colonization and clinical samples were collected during the same period. Results of SCCmec typing and exotoxin profiling for both community MRSA-colonizing strains and CA-MRSA–infecting strains are shown in Table 2. Of 74 colonizing strains that did not have the PVL locus, most (94.6%) had SCCmec IV, and only 1 (1.35%) had SCCmec VT. Conversely, irrespective of origin, PVL-positive MRSA strains were more likely to have SCCmec VT than were the PVL-negative MRSA-colonizing strains (p<0.001). Regarding the exotoxin profiles, the most frequently encoded toxin gene among PVL-positive MRSA isolates was seb (97.9%). PVL-positive MRSA-colonizing strains and CA-MRSA–infecting strains were more likely to have genes that encoded SEB than were PVL-negative CA-MRSA–colonizing strains (p = 0.006). Genes for SEG/SEI were found only in PVL-negative MRSA-colonizing strains (p<0.001).
Figure
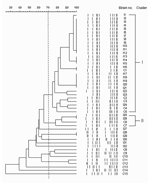
Figure. Pulsed-field gel electrophoresis patterns and phylogenetic tree of 47 methicillin-resistant Staphylococcus aureus (MRSA) isolates with Panton-Valentine leukocidin (PVL) genes. Banding patterns were digitalized and analyzed with Molecular Analyst Fingerprinting, Fingerprinting Plus,...
Diverse pulsotypes were found among the 47 PVL-positive MRSA strains subjected to PFGE typing (Figure). Two clusters that included 33 (70.2%) isolates were distinguished at the 70% similarity level. None of the isolates were linked on review of epidemiologic data derived from medical records. Except for 1 colonizing isolate (C7) and 2 infecting isolates (I22 and I23) that carried SCCmec IV, the other 30 (90.9%) isolates from clusters I and II had SCCmec VT.
This is the first epidemiologic study of PVL-positive S. aureus in Taiwan. The prevalence of PVL-positive S. aureus among isolates collected from various types of staphylococcal infections was 56.9%; previous surveys reported rates of <5% to 12.4% (13–15). This higher prevalence is probably related to the greater proportion of pediatric patients with cutaneous infections. The frequencies of infections associated with these organisms were similar to those in previous studies (2,13,14). Overall, the outcomes of infections with PVL-positive strains were excellent and comparable to that of PVL-negative strains. Only 1 death occurred.
Four factors that support the relationship between nasal strains of PVL-positive MRSA and community-acquired disease. First, both PVL-positive MRSA-colonizing strains and CA-MRSA–infecting strains had consistent antibiograms. Second, most PVL-positive MRSA-colonizing strains and CA-MRSA–infecting strains had SCCmec VT, whereas a high prevalence of SCCmec IV was found among PVL-negative MRSA-colonizing strains. Third, PVL-positive MRSA-colonizing strains and CA-MRSA–infecting strains had consistent exotoxin profiles, which differed from those of PVL-negative MRSA-colonizing strains. Fourth, PFGE findings indicate that some PVL-positive MRSA isolates that colonized children who remained asymptomatic were clonally related to clinically isolated CA-MRSA–infecting strains, especially those with SCCmec VT, compared with SCCmec IV (6/8, 75% and 1/7, 14.3%, respectively).
Several study limitations merit consideration. First, our study is a snapshot in time because the molecular epidemiology of CA-MRSA is constantly changing. Second, we were unable to determine risk for infection because children colonized with community PVL-positive MRSA were not followed-up longitudinally. Finally, our results are geographically distinct and may not be generalized to the global population.
Our study showed that PVL genes are carried by a large number of S. aureus isolates, especially among those causing disease. We provide evidence that links community PVL-positive MRSA-colonizing strains to CA-MRSA–infecting strains from various types of staphylococcal infection.
Dr Lo is a graduate student at the Graduate Institute of Medical Sciences of National Defense Medical Center in Taipei, Taiwan. His research interests include the molecular epidemiology and molecular biology of CA-MRSA infections.
Acknowledgments
We thank L.K. Siu for laboratory support and Shu-Ying Tsai for assistance with and maintenance of the MRSA database.
This study was supported by the National Science Council (grant NSC94-2314-B-016-029) and the Tri-Service General Hospital (grant TSGH-C94-12).
References
- Prevost G, Mourey L, Colin DA, Menestrina G. Staphylococcal pore-forming toxins. Curr Top Microbiol Immunol. 2001;257:53–83.PubMedGoogle Scholar
- Lina G, Piémont Y, Godail-Gamot F, Bes M, Peter MO, Gauduchon V, Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis. 1999;29:1128–32. DOIPubMedGoogle Scholar
- Vandenesch F, Naimi T, Enright MC, Lina G, Nimmo GR, Heffernan H, Community-acquired methicillin-resistant Staphylococcus aureus carrying Panton-Valentine leukocidin genes: worldwide emergence. Emerg Infect Dis. 2003;9:978–84.PubMedGoogle Scholar
- Wang CC, Lo WT, Chu ML, Siu LK. Epidemiological typing of community-acquired methicillin-resistant Staphylococcus aureus isolates from children in Taiwan. Clin Infect Dis. 2004;39:481–7. DOIPubMedGoogle Scholar
- Pan ES, Diep BA, Charlebois ED, Auerswald C, Carleton HA, Sensabaugh GF, Population dynamics of nasal strains of methicillin-resistant Staphylococcus aureus and their relation to community-associated disease activity. J Infect Dis. 2005;192:811–8. DOIPubMedGoogle Scholar
- Hussain FM, Boyle-Vavra S, Bethel CD, Daum RS. Current trends in community-acquired methicillin-resistant Staphylococcus aureus at a tertiary care pediatric facility. Pediatr Infect Dis J. 2000;19:1163–6. DOIPubMedGoogle Scholar
- National Committee for Clinical Laboratory Standards. Methods for disk diffusion: approved standard M2–A8: performance standards for antimicrobial disk susceptibility tests. Wayne (PA): The Committee; 2003.
- National Committee for Clinical Laboratory Standards. Performance standards for antimicrobial susceptibility testing: 14th informational supplement. NCCLS document M100–S14. Wayne (PA): The Committee; 2004.
- Oliveira DC, de Lencastre H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2002;46:2155–61. DOIPubMedGoogle Scholar
- Boyle-Vavra S, Ereshefsky B, Wang CC, Daum RS. Successful multiresistant community-associated methicillin-resistant Staphylococcus aureus lineage from Taipei, Taiwan, that carries either the novel staphylococcal chromosome cassette mec (SCCmec) type VT or SCCmec type IV. J Clin Microbiol. 2005;43:4719–30. DOIPubMedGoogle Scholar
- Jarraud S, Cozon G, Vandenesch F, Bes M, Etienne J, Lina G. Involvement of enterotoxins G and I in staphylococcal toxic shock syndrome and staphylococcal scarlet fever. J Clin Microbiol. 1999;37:2446–9.PubMedGoogle Scholar
- McDougal LK, Steward CD, Killgore GE, Chaitram JM, McAllister SK, Tenover FC. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: establishing a national database. J Clin Microbiol. 2003;41:5113–20. DOIPubMedGoogle Scholar
- Prevost G, Couppie P, Prevost P, Gayet S, Petiau P, Cribier B, Epidemiological data on Staphylococcus aureus strains producing synergohymenotropic toxins. J Med Microbiol. 1995;42:237–45. DOIPubMedGoogle Scholar
- Holmes A, Ganner M, McGuane S, Pitt TL, Cookson BD, Kearns AM. Staphylococcus aureus isolates carrying Panton-Valentine leukocidin genes in England and Wales: frequency, characterization, and association with clinical disease. J Clin Microbiol. 2005;43:2384–90. DOIPubMedGoogle Scholar
- Yamasaki O, Kaneko J, Morizane S, Akiyama H, Arata J, Narita S, The association between Staphylococcus aureus strains carrying Panton-Valentine leukocidin genes and the development of deep-seated follicular infection. Clin Infect Dis. 2005;40:381–5. DOIPubMedGoogle Scholar
Figure
Tables
Cite This ArticleTable of Contents – Volume 12, Number 8—August 2006
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Chih-Chien Wang, Department of Pediatrics, Tri-Service General Hospital, National Defense Medical Center, No. 325, Cheng-Kung Rd, Section 2, Nei-hu 114, Taipei, Taiwan, Republic of China
Top