Volume 18, Number 1—January 2012
Letter
Hantavirus in Bat, Sierra Leone
To the Editor: Hantaviruses (family Bunyaviridae) are transmitted from rodent reservoirs to humans. These viruses cause life-threatening human diseases: hantavirus cardiopulmonary syndrome in the Americas and hemorrhagic fever with renal syndrome in Asia and Europe (1). Since 2006, indigenous hantaviruses were reported also from Africa. Sangassou virus was found in an African wood mouse (Hylomyscus simus) in Guinea (2). Discovery of newer African hantaviruses, Tanganya virus and recently Azagny virus, was even more surprising because they were found in shrews (3,4).
The detection of hantaviruses in small mammals other than rodents, such as shrews and also moles (4), increasingly raises questions regarding the real hantavirus host range. Bats (order Chiroptera) are already known to harbor a broad variety of emerging pathogens, including other bunyaviruses (5). Their ability to fly and social life history enable efficient pathogen maintenance, evolution, and spread. Therefore, we conducted a study on hantaviruses in bats from Africa.
A total of 525 tissue samples from 417 bats representing 28 genera were tested for the presence of hantavirus RNA. Samples originated from different regions in western and central Africa and were collected during 2009 and early 2011. Total RNA was extracted from tissue samples and reverse transcribed. cDNA was screened by PCR specific for sequences of the large genomic segment across the genus Hantavirus (2).
One sample yielded a product of the expected size and was subjected to cloning and sequencing. The positive sample (MGB/1209) was obtained from 1 of 18 investigated slit-faced bats (family Nycteridae). The animal was trapped at the Magboi River within Gola National Park, Sierra Leone (7°50.194′N, 10°38.626′W), and the identification as Nycteris hispida has been verified with the voucher specimen (RCJF529). Histologic examination of organs of the animal showed no obvious pathologic findings.
Figure
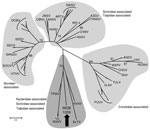
Figure. Maximum-likelihood phylogenetic tree of MGB/1209 virus based on partial large segment sequence (414 nt) and showing the phylogenetic placement of the novel sequence from Nycteris spp. bat compared with hantaviruses associated...
The obtained 414-nt sequence covers a genomic region, which was found to correspond to nt position 2,918–3,332 in the large segment open reading frame of prototypic Hantaan virus. Bioinformatic analysis on the amino acid level showed highest degrees of identity to shrew- and mole-associated hantaviruses (Thottapalayam virus 73.0%, Altai virus 69.7%, Nova and Imjin virus 69.3%). On the basis of tree topology of a maximum-likelihood phylogenetic tree, the sequence does not cluster with rodent-associated hantaviruses but groups with those found in shrews and moles (Figure).
Considering that bats are more closely related to shrews and moles than to rodents (6), a certain genetic similarity of a putative bat-borne hantavirus with shrew- and mole-associated hantaviruses seems reasonable. Notably, shrew-associated Thottapalayam virus (India) and Imjin virus (South Korea) seem to be closer relatives, and African Tanganya virus (Guinea) and Azagny virus (Côte d’Ivoire) are more distantly related. Additional sequence data is needed for more conclusive phylogenetic analyses.
Because the new amino acid sequence is at least 22% divergent from those of other hantaviruses, we conclude that the bat was infected with a newly found hantavirus. We propose the putative name Magboi virus (MGBV) for the new virus because it was detected in an animal captured at the Magboi River in Sierra Leone. The MGBV nucleotide sequence is novel and has not been known or handled before in our laboratory. Before this study, hantavirus nucleic acid was found in lung and kidney tissues of bats from the genera Eptesicus and Rhinolophus in South Korea. However, nucleotide sequencing showed the presence of prototypical Hantaan virus indicating a spillover infection or laboratory contamination (7).
Further screening is necessary to confirm N. hispida as a natural reservoir host of the virus. Although the presented bat-associated sequence is obviously distinct from other hantaviruses, which suggests association with a novel natural host, a spillover infection from another, yet unrecognized host cannot be ruled out. However, detection of the virus exclusively in 1 organ (lung but not in liver, kidney, and spleen; data not shown) suggests a persistent infection that is typically observed in natural hosts of hantaviruses (8).
To date, only a few reports exist on cases of hemorrhagic fever with renal syndrome in Africa (9,10). However, underreporting must be assumed because the symptoms resemble those of many other febrile infections. Moreover, in cases of infections by non–rodent-associated hantaviruses, cross-reactivity with routinely used rodent-borne virus antigens should be limited and may hamper human serodiagnostics (1). The results suggest that bats, which are hosts of many emerging pathogens (5), may act as natural reservoirs for hantavirus. The effect of this virus on public health remains to be determined.
Acknowledgment
This study was supported by Deutsche Forschungsgemeinschaft (grant KR1293/9-1). Work in Sierra Leone was supported by the Gola Forest Project. National and local authorities in Sierra Leone, Liberia, Mali, Senegal, and Republic of Congo kindly granted research and export permits. N.D.W. is supported by an award from the National Institutes of Health Director’s Pioneer Award (grant DP1-OD000370).
References
- Krüger DH, Schonrich G, Klempa B. Human pathogenic hantaviruses and prevention of infection. Hum Vaccin. 2011;7:685–93. DOIPubMedGoogle Scholar
- Klempa B, Fichet-Calvet E, Lecompte E, Auste B, Aniskin V, Meisel H, Hantavirus in African wood mouse, Guinea. Emerg Infect Dis. 2006;12:838–40.PubMedGoogle Scholar
- Klempa B, Fichet-Calvet E, Lecompte E, Auste B, Aniskin V, Meisel H, Novel hantavirus sequences in shrew, Guinea. Emerg Infect Dis. 2007;13:520–2. DOIPubMedGoogle Scholar
- Kang HJ, Kadjo B, Dubey S, Jacquet F, Yanagihara R. Molecular evolution of Azagny virus, a newfound hantavirus harbored by the West African pygmy shrew (Crocidura obscurior) in Cote d’Ivoire. Virol J. 2011;8:373. DOIPubMedGoogle Scholar
- Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T. Bats: important reservoir hosts of emerging viruses. Clin Microbiol Rev. 2006;19:531–45. DOIPubMedGoogle Scholar
- Murphy WJ, Eizirik E, Johnson WE, Zhang YP, Ryder OA, O’Brien SJ. Molecular phylogenetics and the origins of placental mammals. Nature. 2001;409:614–8. DOIPubMedGoogle Scholar
- Jung YT, Kim GR. Genomic characterization of M and S RNA segments of hantaviruses isolated from bats. Acta Virol. 1995;39:231–3.PubMedGoogle Scholar
- Schönrich G, Rang A, Lutteke N, Raftery MJ, Charbonnel N, Ulrich RG. Hantavirus-induced immunity in rodent reservoirs and humans. Immunol Rev. 2008;225:163–89. DOIPubMedGoogle Scholar
- Coulaud X, Chouaib E, Georges AJ, Rollin P, Gonzalez JP. First human case of haemorrhagic fever with renal syndrome in the Central African Republic. Trans R Soc Trop Med Hyg. 1987;81:686. DOIPubMedGoogle Scholar
- Klempa B, Koivogui L, Sylla O, Koulemou K, Auste B, Kruger DH, Serological evidence of human hantavirus infections in Guinea, West Africa. J Infect Dis. 2010;201:1031–4. DOIPubMedGoogle Scholar
Figure
Cite This Article1These authors contributed equally to this article.
Related Links
Table of Contents – Volume 18, Number 1—January 2012
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Detlev H. Kruger, Institute of Medical Virology, Helmut-Ruska-Haus, Charité Medical School, Charitéplatz 1, D-10117 Berlin, Germany
Top