Volume 18, Number 10—October 2012
Dispatch
Influenza A(H1N1)pdm09 Virus in Pigs, Réunion Island
Cite This Article
Citation for Media
Abstract
During 2009, pandemic influenza A(H1N1)pdm09 virus affected humans on Réunion Island. Since then, the virus has sustained circulation among local swine herds, raising concerns about the potential for genetic evolution of the virus and possible retransmission back to humans of variants with increased virulence. Continuous surveillance of A(H1N1)pdm09 infection in pigs is recommended.
Influenza A(H1N1)pdm09 virus, which caused the last influenza pandemic among humans, is a unique reassortant derived from swine influenza viruses of the triple reassortant swine North American lineage and the avian-like swine Eurasian lineage (1). Réunion Island, a tropical French overseas department in the southwestern Indian Ocean, was struck by the influenza pandemic during July–August 2009. The epidemic had a high attack rate in humans (estimated clinically at 12.5% and serologically at 40.0%) (2,3). A(H1N1)pdm09 virus was reported to cause a reverse zoonosis in pigs (4); thus, a long-term (2009–2011) serologic and virologic survey was designed to check for transmission of the virus to pigs on Réunion Island, where the pork industry is a key economic activity and no live pigs have been imported since 1978. At 6-month intervals, a local veterinary surveillance system conducts serologic surveillance for pathogenic swine influenza viruses (H1N1, H1N2, and H3N2) among local herds, and during the last 20 years, none have been detected.
Figure
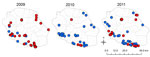
Figure. . Location of farms tested for antibodies against influenza A(H1N1)pdm09 virus in serologic surveys, Réunion Island, 2009–2011. Blue dots, seronegative farms; red dots, seropositive farms.
During a first step (November 2009–February 2010), seroprevalence rates for A(H1N1)pdm09 virus were assessed in 120 breeding pigs (>4 years old) from 57 farms. Blood was obtained from randomly selected pigs at the only slaughterhouse on the island, where pigs are held for <3 hours. We screened the samples for antibodies to influenza A viruses by using the ID Screen Antibody Influenza A kit (ID.vet, Montpellier, France), and titers were determined by using hemagglutination-inhibition (HI) assays (5) against all classical swine influenza viruses and A(H1N1)pdm09 virus (Table 1). Ninety-eight (81.7%; 95% CI 74.7%–88.5%) of the 120 serum samples were positive for A(H1N1)pdm09 virus (HI titers >20); the range of positive titers was 40–640, and 54.2% of the samples expressed high HI titers (160–640). Of the 98 serum samples, 5 reacted at low titer and with only 1 European A (H1N1) swine virus (titer <20), i.e., >4 dilutions lower than for A(H1N1)pdm09 virus, indicating cross reactivity (6). Thus, pigs from 47 (82.4%) of 57 tested farms had been infected by A(H1N1)pdm09 virus; the seroprevalence rate was 81%–100% for pigs on 79.0% of the farms. Farms with affected pigs were located throughout the island (Figure).
In a second step (June 2010, when A(H1N1)pdm09 infection was no longer detected among humans), we tested whether the virus was still circulating among pigs born that year. To obtain nasal swab and blood samples for testing, we randomly selected 390 fattening pigs (25–27 weeks old) at the slaughterhouse; the pigs originated from 45 farms. At the time of sampling, the veterinary surveillance system did not report any clinical signs suggesting virus circulation among herds. However, ≈3.5% of the serum samples (9% of tested farms) contained antibodies to A(H1N1)pdm09 virus (HI titers 20–160). Nasal swab specimens from 6.7% (26/390) of pigs were positive for A(H1N1)pdm09 virus as determined by using a specific real-time reverse transcription PCR (rRT-PCR); the pigs originated from 13 (28.8%) farms (7). Two strains, A/Sw/La Reunion/0164/10 and A/Sw/LaReunion/110348/10, were isolated onto MDCK cell cultures (5).
During July–December 2010, 11 farms reported influenza-like clinical signs in pigs, and proof of A(H1N1)pdm09 virus infection was established on 3 farms (farms A–C). In June 2010, fattening pigs on farm A were seronegative for A(H1N1)pdm09 virus. In July, when acute respiratory disease was reported among pigs, 12 of 39 fattening pigs (18–21 weeks old) sampled on farm A were still seronegative for A(H1N1)pdm09 virus; however, rRT-PCR results were positive for A(H1N1)pdm09 virus. Four weeks later, when pigs had recovered from influenza, only 7.7% (3/39) of sampled pigs on farm A had rRT-PCR results positive for A(H1N1)pdm09 virus, and all 39 were seropositive for the virus. High rates of rRT-PCR positivity were also noted for pigs on farms B (17/30 pigs) and C (6/15 pigs). Two A(H1N1)pdm09 strains (A/Sw/LaReunion/0167/10 and A/Sw/La Reunion/110194/10) were isolated from pigs on farms A and B, respectively.
Four influenza virus strains were isolated from pigs, and all induced a cytopathic effect and displayed hemagglutinating activity on chicken erythrocytes; all 4 were confirmed as A(H1N1)pdm09 virus by specific rRT-PCRs. In addition cross-HI assays (5) revealed that these strains exhibit antigenic relationships with swine influenza A(H1N1) viruses from classical and avian-like lineages, although they reacted most strongly with A(H1N1)pdm09 virus (Table 1). Genome sequencing of these strains showed high (>98%) nucleotide sequence homology to the corresponding genes of A/California/04/09 and 2009 human strains from Réunion Island, suggesting human-to-swine transmission (H. Pascalis, unpub. data).
In a third step (March, July–August, and October 2011), 3 other sampling campaigns were conducted at the slaughterhouse, including 831 fattening pigs from 104 farms. Nasal swab samples for 7 (8.4%) pigs from 3 (2.9%) farms still had rRT-PCR–positive results. However, serologic analyses revealed that pigs on ≈40% of the farms (distributed throughout the island) were seropositive for A(H1N1)pdm09 virus, indicating continuing circulation of the virus in swine herds (Table 2).
Consistent with findings elsewhere (8), our results show that A(H1N1)pdm09 virus has substantially affected swine herds in Réunion Island. Results of our long-term (≈2 years) investigation show that A(H1N1)pdm09 virus has circulated in pigs beyond the 5-week epidemic among humans during the austral winter 2009 (3) and has become a novel enzootic pathogen in Réunion Island.
Several facts may account for the heavy human-to-swine transmission of A(H1N1)pdm09 virus. First, the reassortant pandemic virus contains genomic segments originating from swine influenza viruses established in pigs (1). Second, pigs are highly susceptible to experimental inoculations with A(H1N1)pdm09 virus and support high intraspecies transmissibility (9). Third, the pressure of infection caused by A(H1N1)pdm09 virus among humans in Réunion Island was high but most infections were mild or asymptomatic (3); therefore, people pursued their professional activities, acting as silent spreaders of the virus. Last, pigs on Réunion Island had no history of previous passages of swine influenza viruses; thus, the lack of specific immunity to influenza A viruses would have contributed to the high sensitivity of the pigs to infection, as described (10,11).
Despite serologic proof of large numbers of infected pigs during late 2009–early 2010 in Réunion Island, influenza-like signs were not exhibited and reported until July 2010; this finding was similar to that in New Caledonia (8). In July 2010, several herds showed symptomatic changes in infection that could indicate either a change in virulence of the circulating strain or the intervention of co-infecting pathogens or some other environmental factor(s). Mycoplasma hyopneumoniae and Pasteurella multocida were co-detected on farms with pigs with signs of infection (data not shown). Co-infection with swine influenza virus and these bacteria is known to contribute to severe respiratory disorders among pigs; thus, these bacteria may have enhanced pathogenicity of A(H1N1)pdm09 virus on affected farms (12).
Because specific immunity to A(H1N1)pdm09 virus will decline over time when the virus is no longer circulating among humans, persistence of the virus in an animal reservoir raises concerns about the risk for genetic evolution of the virus and retransmission back to humans of variants with potentially increased virulence. As an example, during the 2011 austral winter, only influenza A(H3N2) and B viruses were recorded (13). Novel reassortant viruses containing genomic segments from A(H1N1)pdm09 and enzootic swine influenza viruses have been isolated in pigs (14). Such a reassortant was responsible for several cases of influenza among humans in 2011 (15); these cases were mild, but other, more virulent pathogenic viruses could emerge. Hence continuous surveillance of A(H1N1)pdm09 infection in pigs is recommended.
Dr Cardinale is in charge of animal and veterinary public health at CRVOI, where he is coordinating a surveillance network on animal diseases in the Indian Ocean entitled “AnimalRisk-OI.” His primary research interests are bacterial and viral zoonoses.
Acknowledgments
We are indebted to Nicolas Rose for helpful discussions.
CRVOI (Le Centre de Recherche et de Veille sur les Maladies Emergentes dans l’Océan Indien) was supported in part by the Ministry of Agriculture through its local representation, DAAF–Réunion (Directorate for Food, Agriculture, and the Forest–Réunion); by FEDER (European Funds for Regional Development) convergence–CPER (Contrat Programme Etat/Region) funds; and by specific funds from the Institut de Microbiologie et Maladies Infectieuses; Institut national de la santé et de la recherché medicalé; Agence Inter-établissements de Recherche pour le Développement; Centre National de la Recherche Scientifique; Institut National de Veille Sanitaire; Agence Nationale de Sécurité Sanitaire, de l’Alimentation, de l’Environnement, et du Travail (ANSES); Institut National de Recherche Agronomique; and Institut Pasteur. Support for work at the French National Reference Laboratory for Swine Influenza was provided by the French Ministry of Agriculture (contracts 2009-175/18277 and 2011-53/2200304774).
References
- Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science. 2009;325:197–201. DOIPubMedGoogle Scholar
- D'Ortenzio E, Renault P, Jaffar-Bandjee MC, Gauzere BA, Lagrange-Xelot M, Fouillet A, A review of the dynamics and severity of the pandemic A(H1N1) influenza virus on Réunion Island, 2009. Clin Microbiol Infect. 2010;16:309–16. DOIPubMedGoogle Scholar
- Dellagi K, Rollot O, Temmam S, Salez N, Guernier V, Pascalis H, Pandemic influenza due to pH1N1/2009 virus: estimation of infection burden in Réunion Island through a prospective serosurvey, austral winter 2009. PLoS ONE. 2011;6:e25738. DOIPubMedGoogle Scholar
- World Organization for Animal Health. World Animal Health Information Database; 2009 [cited 2012 Aug 12]. http://web.oie.int/wahis/public.php?page=weekly_report_index&admin=0
- World Organization for Animal Health. Manual of diagnostic tests and vaccines for terrestrial animals 2010 [cited 2012 Mar 15]. http://www.oie.int/en/international-standard-setting/terrestrial-manual/access-online/
- Kyriakis CS, Olsen CW, Carman S, Brown IH, Brookes SM, Van Doorsselaere J, Serologic cross-reactivity with pandemic (H1N1) 2009 virus in pigs, Europe. Emerg Infect Dis. 2010;16:96–9. DOIPubMedGoogle Scholar
- Pol F, Queguiner S, Gorin S, Deblanc C, Simon G. Validation of commercial real-time RT-PCR kits for detection of influenza A viruses in porcine samples and differentiation of pandemic (H1N1) 2009 virus in pigs. J Virol Methods. 2011;171:241–7. DOIPubMedGoogle Scholar
- Njabo KY, Fuller TL, Chasar A, Pollinger JP, Cattoli G, Terregino C, Pandemic A/H1N1/2009 influenza virus in swine, Cameroon, 2010. Vet Microbiol. 2012;156:189–92. DOIPubMedGoogle Scholar
- Brookes SM, Nunez A, Choudhury B, Matrosovich M, Essen SC, Clifford D, Replication, pathogenesis and transmission of pandemic (H1N1) 2009 virus in non-immune pigs. PLoS ONE. 2010;5:e9068. DOIPubMedGoogle Scholar
- Hofshagen M, Gjerset B, Er C, Tarpai A, Brun E, Dannevig B, Pandemic influenza A(H1N1)v: human to pig transmission in Norway? Euro Surveill. 2009;14:19406.PubMedGoogle Scholar
- Simon G, Hervé S, Rose N, Marchal C. Transmission of pandemic influenza A/H1N1 2009 virus to pigs in New Caledonia, an insular region previously free from swine influenza viruses. In: Proceedings of the 6th International Symposium on Emerging and Re-emerging Pig Diseases; Barcelona, Spain; 2011 Jun 12–15; Poster P191. Barcelona: Centre de Recerca en Sanitat Animal; 2011.
- Fablet C, Marois-Créhan C, Simon G, Grasland B, Jestin A, Kobisch M, Infectious agents associated with respiratory diseases in 125 farrow-to-finish pig herds: a cross-sectional study. Vet Microbiol. 2012;157:152–63. DOIPubMedGoogle Scholar
- Brottet E, Jaffar-Bandjee MC, Valette M, Polycarpe D, Filleul L. Surveillance épidémiologique et virologique de la grippe à la Réunion: un hiver austral 2011 très calme [cited 2012 Mar 10]. http://www.ars.ocean-indien.sante.fr/fileadmin/OceanIndien/Internet/Actualites/BVS_14_decembre_2011.pdf
- Fan X, Zhu H, Zhou B, Smith D, Chen X, Lam T, Emergence and dissemination of a swine H3N2 reassortant influenza virus with 2009 pandemic H1N1 genes in pigs in China. J Virol. 2012;86:2375–8. DOIPubMedGoogle Scholar
- Centers for Disease Control and Prevention. Limited human-to-human transmission of novel influenza A (H3N2) virus—Iowa. MMWR Morb Mortal Wkly Rep. 2011;60:1615–7.PubMedGoogle Scholar
Figure
Tables
Cite This Article1These authors contributed equally to this article.
2These authors contributed equally to this article.
Table of Contents – Volume 18, Number 10—October 2012
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Eric Cardinale, CIRAD-CRVOI-UMR15, Cyroi 2 rue Maxime Rivière, Ste Clotilde 97490, Réunion, France
Top