Influenza A (H3N2) Outbreak, Nepal
Luke T. Daum*†

, Michael Shaw‡, Alexander I. Klimov‡, Linda C. Canas*, Elizabeth A. Macias*, Debra Niemeyer*, James K. Chambers†, Robert Renthal†, Sanjaya K. Shrestha§, Ramesh P. Acharya¶, Shankar P. Huzdar¶, Nirmal Rimal¶, Khin S. Myint#, and Philip Gould*
Author affiliations: *Air Force Institute for Operational Health, Brooks City Base, San Antonio, Texas; †University of Texas, San Antonio, Texas; ‡Centers for Disease Control and Prevention, Atlanta, Georgia; §Walter Reed/Armed Forces Research Institute of Medical Sciences Research Unit-Nepal, Kathmandu, Nepal; ¶Association of Medical Doctors of Asia-Nepal, Kathmandu, Nepal; #US Army Medical Component of the Armed Forces Research Institute of Medical Sciences, Bangkok, Thailand
Main Article
Figure 2
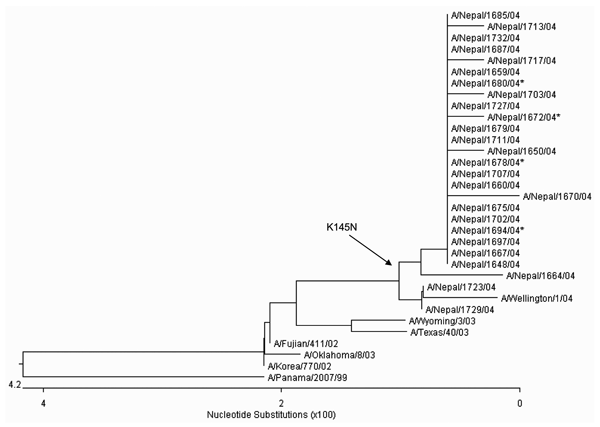
Figure 2. . Unrooted phylogenetic analysis of HA1 hemagglutinin nucleotide sequences from 26 Nepal isolates and H3N2 vaccine and reference strains. The Nepal isolates have drifted from the 2004–2005 A/Fujian/411/03 vaccine strain (and A/Wyoming/03/03 vaccine seed strain) and are genetically equivalent to A/California/7/04, the 2005–2006 Northern Hemisphere vaccine strain. A K145N substitution (branch point indicated by the arrow) was observed in 24 of 26 Nepal isolates and represents a genetic marker for the dominant lineage of H3N2 viruses during the 2004–2005 season. Nucleotide and amino acid sequences for all Nepal isolates are available from GenBank under accession no. AY945263–AY945288. The asterisk indicates isolates from Table 2 that were antigenically distinct from A/Wyoming/303.
Main Article
Page created: April 23, 2012
Page updated: April 23, 2012
Page reviewed: April 23, 2012
The conclusions, findings, and opinions expressed by authors contributing to this journal do not necessarily reflect the official position of the U.S. Department of Health and Human Services, the Public Health Service, the Centers for Disease Control and Prevention, or the authors' affiliated institutions. Use of trade names is for identification only and does not imply endorsement by any of the groups named above.