Volume 12, Number 3—March 2006
Dispatch
Aquariums as Reservoirs for Multidrug-resistant Salmonella Paratyphi B
Cite This Article
Citation for Media
Abstract
Multidrug-resistant Salmonella enterica serovar Paratyphi B dT+ isolates from patients with gastroenteritis were identical with isolates from their home aquariums. Matched isolates had identical phage types, XbaI and IS200 profiles, and Salmonella genomic island 1 (SGI1). Ornamental fish tanks are reservoirs for SGI1-containing S. Paratyphi B dT+.
Figure 1
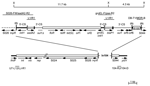
Figure 1. Structure of Salmonella genomic island 1 (SGI1). The SGI1 region of serovar Typhimurium DT104 (GenBank accession no. AF261825) is drawn to scale with In104 above. Different discrete segments are represented by...
Strains of Salmonella enterica serovar Paratyphi B that use d-tartrate as a carbon source (S. Paratyphi B dT+, formerly S. enterica serovar Java) primarily cause gastroenteritis (1). Since the late 1990s, multidrug-resistant S. Paratyphi B dT+ has been increasingly isolated from infected persons in different parts of the world. One type, which is resistant to streptomycin, spectinomycin, trimethoprim, and sulfonamides, carries a chromosomally located class 2 integron with the dfrA1-sat1-aadA1 (Tn7) array of gene cassettes (2). This clone is predominantly associated with poultry and poultry products in Germany and the Netherlands (2,3). Human cases of gastroenteritis caused by S. Paratyphi B dT+ with the resistance phenotype ApCmSmSpSuTc (Ap, ampicillin; Cm, chloramphenicol; Sm, streptomycin; Sp, spectinomycin; Su, sulfonamides; Tc, tetracycline) have also been found in Canada (4), the United Kingdom (5), France (6), and Australia (7), and their incidence is increasing. In most of the studied isolates, the resistance genes blaP1, floR, aadA2, sul1, and tetG are located in a complex class 1 integron recently designated In104 (7) (Figure 1). This integron is located within the Salmonella genomic island 1 (SGI1) that was first identified in S. enterica serovar Typhimurium DT104 strains with the same phenotype (8). However, the source of the SGI1-containing S. Paratyphi B dT+ has not been identified. Whether isolates obtained in different countries are clonally related is also not known.
Although a few epidemiologic studies suggest that antimicrobial drug–susceptible S. Paratyphi B dT+ may be linked to aquacultural practices (9,10), no molecular data confirm this. However, the first reported SGI1-containing S. Paratyphi B dT+ isolate with drug-resistance phenotype ApCmSmSpSuTc was isolated in 1997 from a tropical fish in Singapore (11), raising the possibility that tropical fish and aquariums are a reservoir. The aim of this study was to determine if domestic aquariums are reservoirs for SGI1-containing, multidrug-resistant S. Paratyphi B dT+ that infect humans.
S. Paratyphi B dT+ with the resistance phenotype ApCmSmSpSuTc had been isolated sporadically in various states of Australia since 1997, and initial surveys showed a potential association with ownership of home aquariums (D. Lightfoot, unpub. data). In 2000, multidrug-resistant S. Paratyphi B dT+ with an identical phage type (reaction does not conform [RDNC]), designated here as Aus2, and the same drug-resistance profile (ApCmSmSpSuTc) was isolated from humans with gastroenteritis and from fish tanks in the homes of 2 infected patients (Table). In 2003 and 2004, 13 cases of ApCmSmSpSuTc S. Paratyphi B dT+ were investigated by state and commonwealth health departments, and all were associated with home aquariums containing tropical fish (J. Musto et al., unpub. data). Of these, 11 cases were phage type RDNC Aus3, 1 was phage type 1 var 15, and 1 was phage type 3b var. Water and gravel were collected from the domestic aquariums of 5 patients with RDNC Aus3-type infections, and identical isolates were recovered from each fish tank. Four matched sets of isolates, 2 from 2000 and 2 from 2003, were further examined (Table). One isolate (SRC50) characterized previously (7) was used as a control (Table).
To determine if the resistance phenotype of these strains was due to SGI1 (4,5,7,8,11), polymerase chain reaction (PCR) with primer pairs shown in Figure 1 was used as previously described (7). The left and right junctions of SGI1 with the chromosome and of In104 with SGI1 were present in all cases. Regions containing the gene cassettes were amplified by using standard primers (L1 and R1) in the 5´- and 3´-conserved segments of class 1 integrons. Fragments of 1.0 and 1.2 kb were amplified from all isolates, and digestion of these amplicons with RsaI generated a profile (data not shown) that was indistinguishable from the pattern for the 2 amplicons containing the aadA2 and blaP1 cassettes found in In104 and S. Paratyphi B dT+ isolates SRC49 and SRC50 from 2001 (7). The aadA2 gene cassette was linked to SO26 in the SGI1 backbone, which indicates that it is on the left, as in In104, and the expected 1.8-kb PCR fragment was generated by using primers in groEL and blaP1 (Figure 1), which places the blaP1 cassette on the right. Southern hybridization of Xba I–digested whole-cell DNA with a probe for the floR gene as described previously (7) identified a band of ≈12 kb, which is consistent with an SGI1 structure identical to that reported previously (7,8,11) and the groEL-blaP1 amplicon linked this 12-kb XbaI fragment with the adjacent 4.3-kb XbaI fragment (Figure 1).
Figure 2
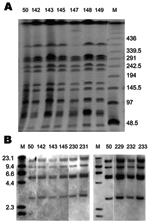
Figure 2. Pulsed-field gel electrophoresis (PFGE) and IS200 profiles of Salmonella enterica serovar Paratyphi B dT+ isolates positive for Salmonella genomic island 1. A) PFGE profiles. Xba I–digested whole-cell DNA was separated by...
To obtain further evidence for the identity of the matched human and fish tank isolates, macrorestriction analyses of XbaI–digested whole-cell DNA by pulsed-field gel electrophoresis (PFGE) were performed as previously described (12). Several studies (3–6,13) suggest that S. Paratyphi B dT+ isolates possess considerable genetic heterogeneity. However, the SGI1-containing isolates appear to be homogeneous. The band patterns for all SGI1-containing S. Paratyphi B dT+ were identical from humans and fish tanks with phage type RDNC Aus3 (Figure 2A) and Aus2 (data not shown). IS200 profiles were also analyzed by hybridization of an IS200 probe with Pst I–digested whole-cell DNA as described elsewhere (6). Again, all strains showed identical profiles (Figure 2B and data not shown) that differed by 1 band from profile IP1 recently described (6). Thus, matched isolates from humans and their fish tanks were indistinguishable from each other.
An unusual observation in this study was that isolates with different phage types showed identical PFGE and IS200 profiles, indicating that they represented a clonal cluster. The control strain SRC50 (RDNC) also displayed the same patterns, demonstrating that it also is a member of the same clone. Thus, variation in phage type (Table) appears to have occurred within a single clone. Variation in phage type has also been reported in other studies of multidrug-resistant S. Paratyphi B dT+ strains (4–6), although a number of related but slightly different XbaI PFGE patterns were observed in those studies. This finding suggests that all multidrug-resistant S. Paratyphi B dT+ found globally have a single origin, but that variations, possibly because of acquisition of other temperate phages or plasmids, have arisen over time. However, direct comparisons of strains from different countries will be needed to confirm this hypothesis.
This is the first definitive report showing that ornamental fish tanks are a reservoir for multidrug-resistant S. Paratyphi B dT+ (ApCmSmSpSuTc phenotype) containing SGI1 that causes severe disease in humans, particularly young children. In addition to containing SGI1, the matched isolates from humans and their fish tanks had the same phage type and the same XbaI macrorestriction digest pattern and IS200 profile. These findings identify home aquariums containing tropical fish as the most important, although not necessarily the only, source of multidrug-resistant S. Paratyphi B dT+. The fact that 12%–14% of Australian households have ornamental fish (14) and as many as 12 million American and 1 million Canadian families own domestic aquariums (9), together with the young age of most affected patients, indicate that multidrug-resistant S. Paratyphi B dT+ in home aquariums is a risk factor for Salmonella infection and thus becomes a public health issue.
Ms Levings is a PhD candidate at the Elizabeth Macarthur Agricultural Institute. Her research interests are the analysis of clustered antimicrobial drug-resistance genes and their spread among members of the Enterobacteriaceae.
Acknowledgments
We thank Linda Falconer for skillful technical assistance.
Renee S. Levings is supported by a grant from the New South Wales Department of Primary Industries and a University of Wollongong Postgraduate Research Award.
References
- Chart H. The pathogenicity of strains of Salmonella Paratyphi B and Salmonella Java. J Appl Microbiol. 2003;94:340–8. DOIPubMedGoogle Scholar
- Van Pelt W, van der Zee H, Wannet WJ, van de Giessen AW, Mevius DJ, Bolder NM, Explosive increase of Salmonella Java in poultry in the Netherlands: consequences for public health. Euro Surveill. 2003;8:31–5.PubMedGoogle Scholar
- Miko A, Guerra B, Schroeter A, Dorn C, Helmuth R. Molecular characterization of multiresistant d-tartrate-positive Salmonella enterica serovar Paratyphi B isolates. J Clin Microbiol. 2002;40:3184–91. DOIPubMedGoogle Scholar
- Mulvey MR, Boyd D, Cloeckaert A, Ahmed R, Ng LK. Emergence of multidrug-resistant Salmonella Paratyphi B dT+, Canada. Emerg Infect Dis. 2004;10:1307–10.PubMedGoogle Scholar
- Threlfall J, Levent B, Hopkins KL, de Pinna E, Ward LR, Brown DJ. Multidrug-resistant Salmonella Java. Emerg Infect Dis. 2005;11:170–1.PubMedGoogle Scholar
- Weill FX, Fabre L, Grandry B, Grimont PA, Casin I. Multiple-antibiotic resistance in Salmonella enterica serotype Paratyphi B isolates collected in France between 2000 and 2003 is due mainly to strains harboring Salmonella genomic islands 1, 1-B, and 1-C. Antimicrob Agents Chemother. 2005;49:2793–801. DOIPubMedGoogle Scholar
- Levings RS, Lightfoot D, Partridge SR, Hall RM, Djordjevic SP. The genomic island SGI1, containing the multiple antibiotic resistance region of Salmonella enterica serovar Typhimurium DT104 or variants of it, is widely distributed in other S. enterica serovars. J Bacteriol. 2005;187:4401–9. DOIPubMedGoogle Scholar
- Boyd D, Peters GA, Cloeckaert A, Boumedine KS, Chaslus-Dancla E, Imberechts H, Complete nucleotide sequence of a 43-kilobase genomic island associated with the multidrug resistance region of Salmonella enterica serovar Typhimurium DT104 and its identification in phage type DT120 and serovar Agona. J Bacteriol. 2001;183:5725–32. DOIPubMedGoogle Scholar
- Gaulin C, Vincent C, Alain L, Ismail J. Outbreak of Salmonella Paratyphi B linked to aquariums in the province of Quebec, 2000. Can Commun Dis Rep. 2002;28:89–93, 96.PubMedGoogle Scholar
- Senanayake SN, Ferson MJ, Botham SJ, Belinfante RT. A child with Salmonella enterica serotype Paratyphi B infection acquired from a fish tank. Med J Aust. 2004;180:250.PubMedGoogle Scholar
- Meunier D, Boyd D, Mulvey MR, Baucheron S, Mammina C, Nastasi A, Salmonella enterica serotype Typhimurium DT104 antibiotic resistance genomic island I in serotype Paratyphi B. Emerg Infect Dis. 2002;8:430–3. DOIPubMedGoogle Scholar
- Thong KL, Ngeow YF, Altwegg M, Navaratnam P, Pang T. Molecular analysis of Salmonella Enteritidis by pulsed-field gel electrophoresis and ribotyping. J Clin Microbiol. 1995;33:1070–4.PubMedGoogle Scholar
- Goh YL, Yasin R, Puthucheary SD, Koh YT, Lim VK, Taib Z, DNA fingerprinting of human isolates of Salmonella enterica serotype Paratyphi B in Malaysia. J Appl Microbiol. 2003;95:1134–42. DOIPubMedGoogle Scholar
- Lehane L, Rawlin GT. Topically acquired bacterial zoonoses from fish: a review. Med J Aust. 2000;173:256–9.PubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 12, Number 3—March 2006
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Steven P. Djordjevic, Microbiology and Immunology Section, New South Wales Department of Primary Industries, Elizabeth Macarthur Agricultural Institute, Private Mail Bag 8, Camden, New South Wales 2570, Australia; fax: 61-2-4640-6384
Top