Volume 14, Number 12—December 2008
Dispatch
Novel Borna Virus in Psittacine Birds with Proventricular Dilatation Disease
Cite This Article
Citation for Media
Abstract
Pyrosequencing of cDNA from brains of parrots with proventricular dilatation disease (PDD), an unexplained fatal inflammatory central, autonomic, and peripheral nervous system disease, showed 2 strains of a novel Borna virus. Real-time PCR confirmed virus presence in brain, proventriculus, and adrenal gland of 3 birds with PDD but not in 4 unaffected birds.
Borna disease virus (BDV) is the causative agent of Borna disease, a meningoencephalitis of horses and sheep in central Europe (1). As the prototype and only known member of the family Bornaviridae in the order Mononegavirales (nonsegmented, negative-strand RNA viruses), BDV is atypical in its nuclear localization of transcription, alternative splicing, and differential use of initiation and termination signals. Sequence analysis of isolates obtained from various species over several decades has shown remarkable sequence conservation; only 2 genotypes are known. The virus is highly neurotropic and infects the central, peripheral, and autonomic nervous systems. Although ungulates remain the best known natural host, the introduction of sensitive molecular and serologic assays enabled by subtractive cloning of the BDV genome facilitated surveys that indicated wider geographic and species distribution (1). Experimental infections are described in a wide variety of vertebrates including chickens, quails, rats, rabbits, cats, shrews, and nonhuman primates; manifestations of disease range from fatal meningoencephalitis to subtle behavioral alterations or asymptomatic persistent infection (2). Intestinal colic is frequently observed in infected ungulates (2,3). An outbreak of neurologic disease in farmed ostriches in Israel has been attributed to BDV (4). BDV nucleic acids have been reported in feces of wild mallards and jackdaws in Sweden (5).
Proventricular dilatation disease (PDD), also known as proventricular dilatation syndrome or macaw wasting disease, is a disorder of birds wherein inflammation of the central, peripheral, and autonomic nervous systems is associated with gastrointestinal dysfunction and neurologic signs that may include ataxia and seizures (6). Although a presumptive diagnosis can be achieved through imaging studies and biopsy, for most animals, definitive diagnosis is made postmortem, only after detailed histologic analysis indicates the accumulation of lymphocytes in nerves that supply the proventriculus and ventriculus, in their associated ganglia, and in brain. Features consistent with PDD have been reported in >50 avian species; however, PDD is most commonly described in exotic companion birds such as macaws and parrots. Whether this reflects better case ascertainment or factors that influence exposure or susceptibility is unclear. An infectious basis is supported by the observation that disease can be transferred to naive birds through inoculation with tissue homogenates or fecal material from affected birds (7). One electron microscopic study showed the presence of spherical, 83-nm particles in macaw embryo cells after inoculation with feces from a diseased macaw; other researchers have described particles in tissue consistent in appearance with adenoviruses or paramyxoviruses. Whether any of these agents can be implicated in the pathogenesis of PDD is unknown.
Figure 1

Figure 1. Conservation of genome organization, regulatory sequences, and protein domains of Borna disease virus (BDV) in novel strains from parrots 1034, 1322, and 1367. N, nucleoprotein; P, phosphoprotein; X, X protein; M,...
Figure 2
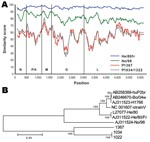
Figure 2. A) Similarity plot between Borna disease virus (BDV) prototype strain V nucleotide sequence and those of characterized BDV strains He/80fr and No/98 compared with novel strains 1367 and 1034/1322. Gene regions...
On a quest for the causative agent of PDD, we investigated 3 birds with a PDD diagnosis based on clinical history and histologic criteria (Table 1). RNA was extracted from brains, pooled and randomly amplified for unbiased high-throughput sequencing (8), yielding 96,698 reads, ranging from 40 nt to 353 nt. After implementation of algorithms for vertebrate sequence subtraction and contiguous fragment assembly, GenBank searches using BLAST (http://blast.ncbi.nlm.nih.gov/blast.cgi) indicated a relation to BDV for a total of 11 contigs covering ≈1.1 kb of sequence distributed in 6 clusters throughout the N (230 nt), P (450 nt), G (250 nt), and L (120, 80, and 250 nt) genes (Figure 1). Divergent sequences between multiple overlapping contigs in these 6 regions indicated at least 2 different strains. Analysis of ≈5.5 kb of genomic sequence generated by standard PCR by using primers (sequence available upon request) based on the identified sequence fragments confirmed 2 strains, one from bird 1367, another from birds 1034 and 1322 (GenBank accession nos. FJ169440 and FJ169441, respectively) and indicated conservation of the unique genome organization that is characteristic for the family Bornaviridae (Figure 1). Sequence divergence between the avian strains (86% identity at nucleotide level) is similar to that observed between the most divergent strains isolated from ungulates (84% identity). Pairwise comparison of the avian strains with these 2 ungulate isolates that represent the 2 previously known genotypes of BDV, strain V, NC_001607 (9), and No/98, AJ311524 (10), indicated <70% sequence conservation at the nucleotide level and <80% at the overall amino acid level (Figure 2, Table 2). These data are compatible with the avian strains representing a new species.
Primers and probes for quantitative real-time PCR were selected in the amino-terminal region of the phosphoprotein (P) gene matching P sequences for strain 1367 (set 1367, forward: 5′-AGAAGACCCGCTGACAGCA-3′, reverse: 5′-AAGCTTCTCGACGGGAACAG-3′, probe: 6FAM-5′-TCGTGGGGACCTCGATCTCACTCG-3′-TMR) or strain 1034/1322 (set 1034–1322, forward: 5′-CAGACAGCACGTCGAGTGAGA-3′, reverse: 5′-AGTTAGGGCCTCCCTGGGTAT-3′, probe: 6FAM-5′-AGGTCCCCGCGAAGGAAGCGA-3′-TMR); the diagnostic assay has been made available through ProMed mail (www.promedmail.org, archive no. 20080726.2287). Real-time PCR showed levels of viral RNA exceeding 103 copies in all tissues tested from birds with PDD but not in control birds. In 2 birds (1034 and 1322) virus load was higher in the brain than in the proventriculus and gizzard or adrenal gland; in 1 bird (1367), the load was lower in the brain than in the adrenals or proventriculus (Table 1).
Western immunoblot and nondenaturing dot blot experiments were pursued by using brain, proventriculus, and adrenal homogenates of the 3 animals that were quantitated by real-time PCR, and 2 rabbit polyclonal antibodies raised against recombinant BDV strain V nucleoprotein (N) or P, as well as immune sera from BDV He/80–infected rats. Positive and negative controls were BDV He/80–infected rat brain homogenate and uninfected rat brain homogenate, respectively. A strong signal was obtained with positive control material in western immunoblots and dot blots that used either antisera to recombinant viral proteins or rat immune sera; no signal was obtained with bird homogenates or uninfected rat brain homogenate.
We have not yet determined whether purified virus induces PDD or an adaptive immune response occurs in association with disease. Nonetheless, given what is known about BDV pathogenesis with other strains in other hosts (11), the bornaviruses identified in these birds must be considered a biologically plausible candidate causative agent. Infection, lymphocyte infiltration, and dysfunction of the central, peripheral, and autonomic nervous system are common to PDD as well as to classical Borna disease in natural disease and experimental models (2,12,13). Proteins and antisera we have used for 2 decades for BDV diagnostics failed to detect this virus in our PCR-positive birds. Thus, it will be important to revisit epidemiologic surveys that we, and others, have undertaken to investigate the role of bornaviruses in human disease (14). From a personal perspective, we are intrigued that whereas molecular discovery of the first BDV in the late 1980s required an investment of 2 years in subtractive cloning (15), high-throughput sequencing, bioinformatics, and sequence databases enabled discovery of these 2 strains in 2 weeks.
Dr Honkavuori is a postdoctoral research scientist at the Center for Infection and Immunity in the Mailman School of Public Health of Columbia University. Her research interests include the identification and characterization of novel pathogens as causative agents of disease.
Acknowledgments
This work was supported by National Institutes of Health awards AI062705, AI051292, HL083850, and AI57158 (Northeast Biodefense Center – Lipkin).
During review of this manuscript similar findings were independently reported by Kistler AL, Gancz A, Clubb S, Skewes-Cox P, Fischer K, Sorber K, et al. Recovery of divergent avian bornaviruses from cases of proventricular dilatation disease: identification of a candidate etiologic agent. Virol J. 2008;5:88.
References
- Lipkin WI, Briese T. Bornaviridae. In: Knipe DM, Howley RM, editors. Virology. 5th ed. Philadelphia: Lippincott, Williams & Wilkins; 2007. p. 1829–51.
- Ludwig H, Kraft W, Kao M, Gosztonyi G, Dahme E, Krey H. Borna virus infection (Borna disease) in naturally and experimentally infected animals: its significance for research and practice [in German]. Tierarztl Prax. 1985;13:421–53.PubMedGoogle Scholar
- Zwick W. Bornasche Krankheit und Enzephalomyelitis der Tiere. In: Gildemeister E, Haagen E, Waldmann O, editors. Handbuch der Viruskrankheiten, 2 Band. Jena (Germany): G. Fischer; 1939. p. 254–354.
- Malkinson M, Weisman Y, Ashash E, Bode L, Ludwig H. Borna disease in ostriches. Vet Rec. 1993;133:304.PubMedGoogle Scholar
- Berg M, Johansson M, Montell H, Berg AL. Wild birds as a possible natural reservoir of Borna disease virus. Epidemiol Infect. 2001;127:173–8. DOIPubMedGoogle Scholar
- Gregory CR, Ritchie BW, Latimer KS, Steffens WL, Pesti D, Campagnoli R, Progress in understanding proventricular dilatation disease. Proceedings of the International Aviculturists Society, 1998 Mar 4–8; Orlando, Florida; 1998.
- Gough RE, Drury SE, Harcourt-Brown NH. Virus-like particles associated with macaw wasting disease. Vet Rec. 1996;139:24.PubMedGoogle Scholar
- Cox-Foster DL, Conlan S, Holmes EC, Palacios G, Evans JD, Moran NA, A metagenomic survey of microbes in honey bee colony collapse disorder. Science. 2007;318:283–7. DOIPubMedGoogle Scholar
- Briese T, Schneemann A, Lewis AJ, Park YS, Kim S, Ludwig H, Genomic organization of Borna disease virus. Proc Natl Acad Sci U S A. 1994;91:4362–6. DOIPubMedGoogle Scholar
- Nowotny N, Kolodziejek J, Jehle CO, Suchy A, Staeheli P, Schwemmle M. Isolation and characterization of a new subtype of Borna disease virus. J Virol. 2000;74:5655–8. DOIPubMedGoogle Scholar
- Ludwig H, Bode L, Gosztonyi G. Borna disease: a persistent virus infection of the central nervous system. Prog Med Virol. 1988;35:107–51.PubMedGoogle Scholar
- Stitz L, Bilzer T, Richt JA, Rott R. Pathogenesis of Borna disease. Arch Virol Suppl. 1993;7:135–51.PubMedGoogle Scholar
- Briese T, Hornig M, Lipkin WI. Bornavirus immunopathogenesis in rodents: models for human neurological diseases. J Neurovirol. 1999;5:604–12. DOIPubMedGoogle Scholar
- Lipkin WI, Hornig M, Briese T. Borna disease virus and neuropsychiatric disease–a reappraisal. Trends Microbiol. 2001;9:295–8. DOIPubMedGoogle Scholar
- Lipkin WI, Travis GH, Carbone KM, Wilson MC. Isolation and characterization of Borna disease agent cDNA clones. Proc Natl Acad Sci U S A. 1990;87:4184–8. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 14, Number 12—December 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Thomas Briese, Center for Infection and Immunity, Mailman School of Public Health, Columbia University, 722 W 168th St, 18th Floor, New York, NY 10032, USA
Top