Volume 16, Number 12—December 2010
Dispatch
Increasing Contact with Hepatitis E Virus in Red Deer, Spain
Cite This Article
Citation for Media
Abstract
To describe the epidemiology of hepatitis E virus (HEV) in red deer in mainland Spain, we tested red deer for HEV RNA and antibodies. Overall, 10.4% and 13.6% of serum samples were positive by ELISA and reverse transcription–PCR, respectively. The increasing prevalence suggests a potential risk for humans.
Hepatitis E virus (HEV) is the only member of the Hepeviridae family (1). Four major genotypes of HEV have been recognized: genotypes 1 and 2 are restricted to humans and associated with epidemics in developing countries; genotypes 3 and 4 are zoonotic in developing and industrialized countries. Wild and domestic animals are being identified as potential HEV reservoirs (1–3).
Studies on wild sika deer (Cervus nippon) have detected low prevalence rates for HEV, which suggests that sika deer are accidental hosts for the virus (4,5), despite the transmission link discovered between them and HEV in Japan (3) that raised awareness of the possibility that game animals transmit HEV (2). In Europe, information about HEV infection in wild ruminants is limited to reports suggesting that roe deer (Capreolus capreolus) and red deer (Cervus elaphus) can act as HEV hosts (6–8). Except for these limited studies, no large-scale surveys have been conducted of HEV epidemiology in wild cervids. In Spain, the relatively high HEV seroprevalence detected in domestic pigs and wild boar suggests that HEV infection is probably widespread (9).
Red deer density, distribution, and hunting harvest are increasing throughout Europe (10). In Spain, the high densities recorded (11) indicate that red deer are an important source of game meat. This scenario emphasizes the need for a better understanding of the epidemiology of HEV in game populations in Spain.
Our goals were to describe the epidemiology and time trends of HEV in red deer in peninsular (mainland) Spain by serologic testing and PCR. On the basis of previous results on wild boar (9), we hypothesized that red deer would show widespread contact with HEV in Spain.
Figure
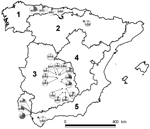
Figure. The 5 peninsular bioregions (nos. 1–5) and the 21 sampling sites, Spain. Pie charts indicate local prevalence (in gray). Numbers indicate positive animals/sampled animals. Broken line borders indicate open sites; solid...
Serum samples from 968 Iberian red deer were collected during 2000–2009. These samples came from hunter-harvested red deer in 21 wild or semifree ranging populations (892 deer) and from 2 farms (76 deer). Sampling sites were representative of a variety of habitats and climates, which can be simplified into 5 different bioregions (Figure) (12). Sampling sites were grouped into 7 areas and 2 red deer farms (Table; Figure). Sex and age of deer were recorded. Management conditions of red deer were classified as open (no fencing and no management, 9 sites), fenced (fencing and artificial feeding, 12 sites), and farmed (livestock-like management, 2 farms). To analyze time trends, we classified samples collected during 2000–2005 as time 1 and those collected during 2006–2009 as time 2. Only sites where sampling occurred in both periods and with comparable sampling sizes were included in the time-trend analysis.
Serum samples were tested for HEV immunoglobulin (Ig) G by using ELISA as described (4,13), except for including protein G horseradish peroxidase (Sigma Chemical, St. Louis, MO, USA) as a conjugate, as in previous studies of red deer (12). Anti-HEV–positive serum was obtained from domestic swine that were positive for HEV by ELISA and reverse transcription–PCR (RT-PCR). Anti-HEV–negative serum was obtained from previous studies (14) and negative controls from HEV-negative cattle (13). Results were expressed as the percentage of optical density (% OD) by using the formula [% OD = 100 × sample OD/sum of negative controls OD]. Serum samples with % OD values >100% were considered positive.
For the RT-PCR, 81 serum samples were randomly selected and analyzed. Viral RNA was extracted from 150 mL of serum with Nucleospin RNA virus kit (Macherey-Nagel, Düren, Germany) by following the manufacturer’s instructions. HEV was detected by using a seminested RT-PCR as described (14). In each run, negative and positive controls were added.
Eight HEV RT-PCR–positive samples were sequenced. HEV sequences were identified by using the BLAST algorithm (www.ncbi.org) against HEV sequences available in GenBank (on January 25, 2010). Sequences were deposited in the GenBank database under accession nos. HM113373 and HM113374.
Sterne exact method was used to estimate apparent prevalence confidence intervals (CIs). χ2 tests were used to analyze the association of age, sex, sampling site, and management conditions with serologic and RT-PCR results. Association between seropositivity and HEV RNA in the serum was also analyzed by using Pearson χ2 test. Differences were considered statistically significant at p<0.05.
Overall, 101 (10.4%, 95% CI 8.6–12.5) serum samples were positive for IgG (Table). HEV seroprevalence did not differ significantly between sex (χ2 = 0.894, 1 df, p>0.05) and age classes (χ2 = 12.436, 3 df, p>0.05). Seroprevalence in time 2 (12.2%, 95% CI 9.8–15.0) was significantly higher than seroprevalence in time 1 (7.5%, 95% CI 5.1–10.8) (χ2 = 5.181, 1 df, p<0.05). Local IgG seroprevalences ranged from 0 (95% CI 0–20.8) to 31.4% (95% CI 21.3–43.5; Figure). IgG seroprevalence differed significantly by management types (χ2 6.876, 2 df, p<0.05), with higher values in open (14.9%, 95% CI 11.3–19.4) than in fenced (9.1%, 95% CI 7.0–11.7) and farmed (2.6%, 95% CI 0.5–9.0) areas.
Eleven (13.6%, 95% CI 7.4–22.7) of 81 samples were RT-PCR positive. Local viral RNA prevalence ranged from 4.5% (95% CI 2.4–22.21 to 38.5% (95% CI 16.6–65.8; Table). HEV prevalence did not differ significantly by geographic area and management type.
Sequence analysis indicated that all deer sequences from this study belonged to genotype 3. Seven samples belonging to sequence HM113374 shared 99% nucleotide identity with domestic swine strains from Spain. One sample, sequence HM113373, showed similarity (91%) with a strain from acute hepatitis E in a person in Marseille, France, according to GenBank.
Our findings of HEV infection confirm that HEV circulates actively among red deer in the Iberian Peninsula, as described for wild boar (9). Red deer can be infected with HEV (7,8), and the results of our large serosurvey in this species in Europe show an increasing prevalence trend in the last decade.
de Deus et al. found higher IgG seroprevalences in estates with higher wild boar densities (9). However, in the present study, mean seroprevalence rates were lowest in red deer farms, where densities were the highest and red deer had no contact with wild boar or domestic swine. In contrast, the highest seroprevalence rates were reported in open areas where contact with suids may have occurred. However, wild boar densities also are high in fenced hunting estates (15), and HEV antibody prevalence rate was intermediate in deer from these sites. These differences could indicate that red deer may need a source of infection and thus act as spillover hosts more frequently than as true reservoirs.
Presence of HEV RNA in 13% of deer serum implies that deer represent a risk for zoonotic transmission, and consequently, handling of live animals and carcasses is a risk activity. Red deer are infected with HEV at lower rates than are wild boar and domestic pigs but may act as a potential source of HEV infection in humans. Further studies are needed to fully elucidate the epidemiology of HEV in wildlife and the foodborne zoonotic transmission risks.
Ms Boadella is a PhD student at the Instituto de Investigación en Recursos Cinegéticos. Her research deals with various aspects of shared wildlife diseases, including time trends and risk factors in diseases of wild ungulates.
Acknowledgments
We thank Bibiana Peralta for providing the HEV antigen protein for ELISA and RT-PCR–positive controls. We also thank our many colleagues at Instituto de Investigación en Recursos Cinegéticos who helped in field and laboratory work.
This study was financed by FISCAM PI-2007/56. The study also benefited from agreements of IREC with MARM-OAPN, Castilla–La Mancha and Principado de Asturias. Funding for sampling was also granted by AGL2008-03875. CReSA thanks CONSOLIDER (CSD-20060007).
References
- Panda SK, Thakral D, Rehman S. Hepatitis E virus. Rev Med Virol. 2007;17:151–80. DOIPubMedGoogle Scholar
- Teo CG. Much meat, much malady: changing perceptions of the epidemiology of hepatitis E. Clin Microbiol Infect. 2010;16:24–32. DOIPubMedGoogle Scholar
- Tei S, Kitajima N, Takahashi K, Mishiro S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet. 2003;362:371–3. DOIPubMedGoogle Scholar
- Matsuura Y, Suzuki M, Yoshimatsu K, Arikawa J, Takashima I, Yokoyama M, Prevalence of antibody to hepatitis E virus among wild sika deer, Cervus nippon, in Japan. Arch Virol. 2007;152:1375–81. DOIPubMedGoogle Scholar
- Yu C, Zimmerman C, Stone R, Engle RE, Elkins W, Nardone GA, Using improved technology for filter paper-based blood collection to survey wild sika deer for antibodies to hepatitis E virus. J Virol Methods. 2007;142:143–50. DOIPubMedGoogle Scholar
- Reuter G, Fodor D, Forgách P, Kátai A, Szucs G. Characterization and zoonotic potential of endemic hepatitis E virus (HEV) strains in humans and animals in Hungary. J Clin Virol. 2009;44:277–81. DOIPubMedGoogle Scholar
- Forgách P, Nowotny N, Erdélyi K, Boncz A, Zentai J, Szucs G, Detection of hepatitis E virus in samples of animal origin collected in Hungary. Vet Microbiol. 2010;143:106–16. DOIPubMedGoogle Scholar
- Rutjes SA, Lodder-Verschoor F, Lodder WJ, van der Giessen J, Reesink H, Bouwknegt M, Seroprevalence and molecular detection of hepatitis E virus in wild boar and red deer in the Netherlands. J Virol Methods. 2010;168:197–206. DOIPubMedGoogle Scholar
- de Deus N, Peralta B, Pina S, Allepuz A, Mateu E, Vidal D, Epidemiological study of hepatitis E virus infection in European wild boars (Sus scrofa) in Spain. Vet Microbiol. 2008;129:163–70. DOIPubMedGoogle Scholar
- Milner JM, Bonenfant C, Myd JM, Csányi S, Stenseth NC. Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors. J Appl Ecol. 2006;43:721–34. DOIGoogle Scholar
- Acevedo P, Ruiz-Fons F, Vicente J, Reyes-Garcia AR, Alzaga V, Gortazar C. Estimating red deer abundance in a wide range of management situations in Mediterranean habitats. J Zool (Lond). 2008;276:37–47. DOIGoogle Scholar
- Muñoz PM, Boadella M, Arnal M, de Miguel MJ, Revilla M, Martínez D, Spatial distribution and risk factors of brucellosis in Iberian wild ungulates. BMC Infect Dis. 2010;10:46. DOIPubMedGoogle Scholar
- Peralta B, Casas M, De Deus N, Martín M, Ortuño A, Pérez-Martín E, Anti-HEV antibodies in domestic animal species and rodents from Spain using a genotype 3-based ELISA. Vet Microbiol. 2009;137:66–73. DOIPubMedGoogle Scholar
- de Deus N, Seminati C, Pina S, Mateu E, Martin M, Segales J. Detection of hepatitis E virus in liver, mesenteric lymph node, serum, bile and faeces of naturally infected pigs affected by different pathological conditions. Vet Microbiol. 2007;119:105–14. DOIPubMedGoogle Scholar
- Acevedo P, Vicente J, Hofle U, Cassinello J, Ruiz-Fons F, Gortazar C. Estimation of European wild boar relative abundance and aggregation: a novel method in epidemiological risk assessment. Epidemiol Infect. 2007;135:519–27. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleTable of Contents – Volume 16, Number 12—December 2010
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Mariana Boadella, Instituto de Investigación en Recursos Cinegéticos, Consejo Superior de Investigaciones Científicas, CSIC-Universidad de Castilla–La Mancha, Junta de Comunidades de Castilla–La Mancha, Ronda de Toledo s/n, 13005 Ciudad Real, Spain
Top