Volume 16, Number 9—September 2010
Research
Cercarial Dermatitis Transmitted by Exotic Marine Snail
Abstract
Cercarial dermatitis (swimmer’s itch) is caused by the penetration of human skin by cercariae of schistosome parasites that develop in and are released from snail hosts. Cercarial dermatitis is frequently acquired in freshwater habitats, and less commonly in marine or estuarine waters. To investigate reports of a dermatitis outbreak in San Francisco Bay, California, we surveyed local snails for schistosome infections during 2005–2008. We found schistosomes only in Haminoea japonica, an Asian snail first reported in San Francisco Bay in 1999. Genetic markers place this schistosome within a large clade of avian schistosomes, but do not match any species for which there are genetic data. It is the second known schistosome species to cause dermatitis in western North American coastal waters; these species are transmitted by exotic snails. Introduction of exotic hosts can support unexpected emergence of an unknown parasite with serious medical or veterinary implications.
One consequence of introduction of exotic species is possible establishment of new host–parasite associations, potentially resulting in emergence of new diseases (1–3). Exotic parasites can be introduced into new locations along with their exotic host species (1,4), sometimes causing extinction of indigenous parasites (5). Newly introduced parasites can extend their host ranges into related indigenous host species (6,7), or exotic hosts may play new roles in the transmission of indigenous parasites (8). Parasites newly supported by these exotic hosts can assume considerable human or animal roles as emerging disease agents (9).
San Francisco Bay has been the site of numerous well-documented introductions of exotic species (10–12). We document an outbreak of human cercarial dermatitis in San Francisco Bay that was related to the recent introduction of an exotic snail, the Japanese bubble snail Haminoea japonica Pilsbury 1895 (Cephalaspidea: Haminoeidae), which serves as the intermediate host of a schistosome that is responsible for the now annual dermatitis outbreaks.
Cercarial dermatitis (swimmer’s itch) is caused by penetration of human skin with cercariae of schistosome parasites; the condition is common and recurrent in freshwater habitats worldwide. Adult schistosomes typically live in mesenteric blood vessels of birds or mammals and produce eggs that pass from the host in feces. The eggs then hatch and release miracidia, which penetrate and develop in an appropriate species of an intermediate snail host. Snail infections culminate in asexual production of numerous cercariae, which are regularly released into the water where they seek to penetrate the skin of a definitive vertebrate host. Penner reported an association between human dermatitis and a marine schistosome (13). He established that Littorina keenae Rosewater 1978 (Hypsogastropoda: Littorinidae) syn. L. planaxis snails collected along the rocky shores of southern California released schistosome cercariae that caused dermatitis in experimentally exposed human volunteers. Documented cases (1,14–21) have been attributed to species of Austrobilharzia Johnston 1917, the adults of which most commonly infect gulls and shorebirds (14).
A cercarial dermatitis outbreak in San Francisco Bay was reported in 1954 (1). The Bureau of Vector Control of the California Department of Health Services identified the cercariae as Austrobilharzia variglandis (Miller and Northrup 1926) collected from the eastern Atlantic mudsnail Ilyanassa obsoleta (Say 1822) (Hypsogastropoda: Nassariidae) syn. Nassarius obsoletus at Robert Crown Memorial Beach in 1955 and 1956 (1). I. obsoleta snails were accidentally introduced into San Francisco Bay in commercial shipments of Atlantic oysters and were observed in the bay in 1907 (10,11). A. variglandis schistosomes had been identified as the cause of cercarial dermatitis in coastal waters in the northeastern United States (14).
After the 1954 outbreak, no additional cases of cercarial dermatitis were reported in San Francisco Bay or elsewhere on the Pacific Coast until the summer of 2001 when 36 cases of cercarial dermatitis were reported at Crescent Beach in Surrey, British Columbia, Canada; 44 cases were reported in 2002 (20). The agent again appeared to A. variglandis schistosomes carried by I. obsoleta snails, which are extremely abundant at Crescent Beach. The snail was accidentally introduced into British Columbia in the oyster trade and was first observed in the region in 1952 (20).
In June 2005, cercarial dermatitis appeared again in San Francisco Bay. The Alameda County Department of Environmental Health received ≈90 reports of skin irritation that occurred after water contact at Robert Crown Memorial Beach where the 1954 outbreak had occurred (The Swimmer Itch Hotline, www.acgov.org/aceh/recreational/beaches.htm). Although it was initially assumed that A. variglandis schistosomes, carried by I. obsoleta snails, were the causative agent, preliminary investigations suggested that the intermediate host involved was another exotic species, Haminoea japonica, from Asia. This snail was observed in San Francisco Bay in 1999 (22) and on the eastern shore of the bay, near Robert Crown Memorial Beach, in the summer of 2003, where it has become extremely abundant (A.N. Cohen, unpub. data). We describe the morphologic and genetic features of cercariae obtained from H. japonica snails and discuss possible transmission pathways of this parasite in San Francisco Bay.
Specimen Collection
Figure 1
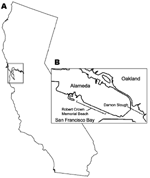
Figure 1. San Francisco Bay area, California, USA (A), and locations where Haminoea japonica snails were obtained (B).
Figure 2
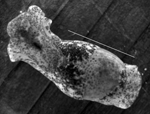
Figure 2. Haminoea japonica snail. Scale bar = 11 µm.
This study was conducted under the University of New Mexico Institutional Animal Care and Use Committee Protocol 07UNM011, Animal Welfare Assurance # A4023-01. Samples of H. japonica snails were collected by hand at low tide from 4 locations (Figure 1): Crab Cove at the northern end of Robert Crown Memorial Beach (37°46′4.18′′N, 122°16′39.55′′W) in 2005–2008; the southern end of this beach (37°45′8.81′′N, 122°14′57.33′′W) in 2006 and 2008; Damon Slough (37°45′14.12′′N, 122°12′21.6′′W) in 2007; and Lake Merritt, a tidal lagoon connected to San Francisco Bay (37°48′22.25′′N, 122°15′22.81′′W) in 2008. All gastropod species observed at Robert Crown Memorial Beach were collected and examined for cercariae. H. japonica (Figure 2), Philine sp. (Cephalaspidea: Philinidae), and I. obsoleta snails (all exotic) were collected on tide flat sediments. The exotic oyster drill Urosalpinx cinerea (Say 1822) (Sorbeoconcha: Muricidae) and the native periwinkle Littorina spp. (L. scutulata Gould 1849 or L. plena Gould 1849) were collected from exposed bulwarks and other hard substrates. In addition, we examined native H. virescens Sowerby 1833 snails collected in August 2005 from Friday Harbor, San Juan Island, Washington.
Snails were isolated singly or in groups of 5 in plastic containers in saline water (20–35 parts per thousand [ppt]) and placed in natural light to induce cercarial shedding. For each species, a subset of the larger snails was then dissected. Cercariae were photographed, and ethanol-preserved specimens were measured and compared with published descriptions of schistosome cercariae.
Gulls are common hosts for schistosomes transmitted by marine snails. We examined 29 gulls of 4 species collected at the Oakland International Airport, ≈7.5 miles southeast of Robert Crown Memorial Beach, as part of the airport’s Wildlife Management Program. For each bird, the mesenteric veins were examined, the intestine was opened, the muscosa was removed, and a sample from the intestinal wall was placed between 2 glass slides and examined for schistosome adults or eggs in the villi (23). Feces were screened for eggs.
Sequencing and Phylogenetic Analysis
DNA was extracted from fresh or ethanol-preserved cercariae, amplified by PCR (Takara Ex Taq; Takara Biomedicals, Otsu, Japan), and sequenced by using published primers (24,25). PCR products were purified on Montage Microcon columns (Millipore, Billerica, MA, USA). Sequencing reactions were performed by using the BigDye Direct Sequencing Kit Version 3.1 (Applied Biosystems, Foster City, CA, USA).
Phylogenetic analyses of schistosomes obtained from H. japonica snails were performed for 2 datasets. The first dataset combined 18S and 28S rRNA sequence data to place our samples of H. japonica within the larger context of the family Schistosomatidae (24). The second dataset, which was composed of part of the internal transcribed spacer region 2, focused on taxa within the schistosome BTGD clade, which includes species of Bilharziella, Trichobilharzia, Gigantobilharzia, and Dendritobilharzia (sensu 24). Phylogenetic analyses with maximum parsimony (MP), maximum likelihood (ML), and minimum evolution (ME) were conducted by using PAUP* version 4.0b10 (26) and Bayesian inference (BI) by using MRBAYES 3 (27). The jModeltest (28) was used to determine the most appropriate nucleotide substitution model for ML and ME analyses.
Parsimony trees were reconstructed by using heuristic searches (300 replicates). Optimal ME and ML trees were constructed from heuristic searches (300 replicates for ME, 10 replicates for ML). Nodal support was estimated by bootstrap (200 replicates) analysis and determined for the MP and ME trees by using heuristic searches. For the ML dataset, the model selected (Akaike information model) was generalized time reversible + proportion invariant + Γ. For BI of the 18S–28S rRNA dataset, the parameters were unlinked: Nst = 6 rates = gamma ngammacat = 4. For both datasets, 4 chains were run simultaneously for 5 × 105 generations; the first 5,000 trees with preasymptotic likelihood scores were discarded as burning, and the retained trees were used to generate 50% majority-rule consensus trees and posterior probabilities.
Life Cycle
Figure 3
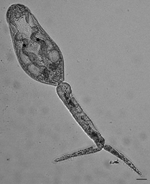
To obtain adult worms for species identification, we experimentally exposed young parakeets and Gallus gallus L. chicks for 30 min to schistosome cercariae from H. japonica snails obtained from San Francisco Bay (Figure 3). Bird hosts were selected according to the method of Leigh (23), who described adult worms derived from cercariae from H. antillarum guadalupensis Sowerby snails in Florida. In experiment 1, eight chicks were exposed by standing each chick in salt water (35 ppt) containing cercariae. In experiment 2, six parakeets were exposed by applying cercariae to their bare abdomens. In experiment 3, eight parakeets were exposed by standing each bird in salt water (35 ppt) containing cercariae.
Specimens Collected
We identified 1 species of schistosome in H. japonica snails that had a prevalence of 1.2% as determined by observation of shedding and 8.7% as determined by dissection (Table 1). No schistosomes were found in other snails examined, including Littorina spp. and I. obsoleta, taxa that are known to host schistosomes in California (1,13). No other trematodes were found in H. japonica snails from San Francisco Bay or H. virescens snails from Washington State. A. variglandis was the only schistosome found in the gulls; 55% had adult worms in their mesenteric veins (Table 1).
Cercariae, most of which were collected by dissection, lay in contact with the surface film of water, where they were mostly inactive except for occasional tail twitching. The cercariae were apharyngeate with pigmented eyespots, a lightly spined body, dorsal and ventral fin folds on the full length of the tail furcae, 5 pairs of flame cells, and 3 pairs of penetration glands (Figure 3). With respect to these behavioral and morphologic features, and on the basis of size, the cercariae most closely resemble those of Gigantobilharzia huttoni (Leigh 1953), the only other schistosome previously collected from haminoeid snails (23,29) (Table 2). These cercariae differ from those of G. huttoni in having fewer pairs of penetration glands (3 pairs instead of 5–6 pairs), but this trait is difficult to discern accurately. In contrast, A. variglandis cercariae have 6 pairs of penetration glands and 6 pairs of flame cells, are larger, and have different proportions than cercariae we found (Table 2). Specimens of the schistosome obtained from H. japonica snails were deposited in the Parasite Division of the Museum of Southwestern Biology at the University of New Mexico (MSB185).
Sequencing and Phylogenetic Analysis
Figure 4
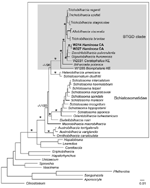
Figure 4. Maximum-likelihood phylogenetic tree based on 18S–28S rRNA sequences of schistosomes. Schistosomatids are indicated in the large box and the Bilharziella, Trichobilharzia, Gigantobilharzia, and Dendritobilharzia (BTGD) clade is indicated in the gray...
Figure 5
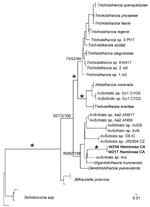
Figure 5. Maximum-likelihood phylogenetic tree based on internal transcribed spacer region 2 sequences of relationships among members of the Bilharziella, Trichobilharzia, Gigantobilharzia, and Dendritobilharzia species clade from this study and unidentified samples of...
Schistosome taxa used in the phylogenetic analyses are shown in the Table A1. Cercariae from H. japonica were distinct from all other available schistosomes (GenBank accession nos. GQ920617–21). These cercariae belong to the BTGD clade, which contains only freshwater schistosomes (Figure 4). Our internal transcribed spacer region 2 dataset includes all reported schistosomes from GenBank that belong to the BTGD clade (Figure 5). Only an ML tree is shown in Figure 5. However, MP, ME, and BI analyses yielded near identical topologies with differences at the tips and at nodes where there is no clade support.
Life Cycle
Three experiments with birds were conducted to obtain adult worms. However, all birds experimentally exposed were negative for schistosome infection.
Cercarial morphology and molecular genetic data for the schistosome from H. japonica indicate that these cercariae are not A. variglandis, a schistosome previously reported in San Francisco Bay, and the only species previously implicated in dermatitis outbreaks on the Pacific Coast. We obtained A. variglandis schistosomes from gulls but not from snails in the San Francisco Bay area. The H. japonica–transmitted schistosome is the second species reported to cause dermatitis after introduction of an exotic snail in California coastal waters. We found this schistosome in an opisthobranch snail in western North America. The only other schistosome known to be obtained from an opisthobranch snail is G. huttoni from H. a. guadalupensis snails obtained in Florida (23,29,32). Except for schistosomes collected from 2 species of Siphonaria (Pulmonata: Siphonariidae) snails (33,34), all other marine schistosomes have been obtained from caenogastropodid snails.
Cercariae from H. japonica closely resemble those of G. huttoni, for which sequence data are not available. DNA sequence data for cercariae from H. japonica did not match with those of any known schistosome species, including the congener G. huronensis Najim 1950. Cercariae from H. japonica did not group with other marine schistosomes, but belong to the BTGD clade that, until now, included only freshwater species that use pulmonate snails as intermediate hosts (Figure 5). The only other known marine schistosomes belong to species of the genera Austrobilharzia and Ornithobilharzia Ohdner 1912, which are distantly related to the BTGD clade (Figure 4). We exposed parakeets and chicks to cercariae from H. japonica snails but were unable to obtain adult worms for comparison with described species.
Schistosomes from H. japonica and those of G. huttoni are probably closely related because they are found in haminoeid snails and have morphologically similar cercariae. However, they differ from all other species in the genus Gigantobilharzia Ohdner 1910 in habitat (salt water rather than fresh water) and snail host (opisthobranchid rather than pulmonate). When appropriate genetic material becomes available, analyses may show that G. huttoni schistosomes and those from H. japonica snails should be placed in a separate genus.
Gigantobilharzia spp. have been reported in several gull species (35). Gulls are common at Robert Crown Memorial Beach, often resting on the beach flats at low tides and sometimes foraging in tide pools that contain H. japonica snails (A.N. Cohen, unpub. data). Gulls are thus a likely host for schistosomes from H. japonica, although we did not find them in gulls at the Oakland Airport near Robert Crown Memorial Beach. Leigh (36) and Kinsella et al. (37) found fragments of adult worms that they identified as Gigantobilharzia sp., and which closely resembled G. huttoni worms, in pelicans in Florida. Small flocks of brown pelicans and, rarely, white pelicans have been observed in shallow water off Robert Crown Memorial Beach, although not on the beach or in tide pools (A.N. Cohen, unpub. data). Pelicans are thus another possible host for schistosomes from H. japonica.
Other birds commonly observed at Robert Crown Memorial Beach include shorebirds that are most common in winter when water temperatures are probably less conducive to cercarial emergence (surface temperatures near this beach are typically 16°C–20°C in summer and 8°C–12°C in winter). Cormorants, grebes, and ducks are found in near-shore waters, and mallard ducks sometimes forage in tide pools. Larger wading birds (snowy and great egrets, and occasionally herons), oystercatchers, and several species of terns are sometimes seen in small numbers foraging on the beach or in shallows. Marine schistosomes have been reported in gulls, ducks, terns, herons, cormorants, and turnstones (14,17,19,23), and Gigantobilharzia spp. have been reported in grebes and cattle egrets (23,35). Thus, various bird species might serve as hosts for schistosomes from H. japonica.
Cercarial dermatitis is commonly acquired in fresh water (38). It is less common in marine or estuarine waters; most cases are reported from the northwestern Atlantic Ocean or Australia. This disease was observed on the Pacific Coast of North America during an outbreak at Robert Crown Memorial Beach in 1954–1956, when cercariae identified as A. variglandis were found in I. obsoleta, an Atlantic snail introduced before 1907. The schistosome was likely introduced with this snail and remained undetected until the 1950s (1). In June 2005, cercarial dermatitis was again reported at Robert Crown Memorial Beach. Initial cases were found among elementary school groups that visited the beach at the end of the academic year. Since 2005, dermatitis has occurred annually at this beach (90 cases in 2005, 3 in 2006, 14 in 2007, and 31 in 2008).
Schistosome cercariae in H. japonica snails, large numbers of these snails at Robert Crown Memorial Beach, and the apparent absence of schistosomes in other common snails at this site indicate that schistosomes from H. japonica are responsible for the recent dermatitis outbreak. H. japonica snails were first seen in California in 1999 (22) and except for the 1954 outbreak attributed to A. variglandis, cercarial dermatitis was not reported in San Francisco Bay until shortly after the arrival of H. japonica.
Figure 6

Figure 6. Cercarial dermatitis contracted in San Francisco Bay, California, USA, by one of the authors (S.V.B.).
The most popular water-contact activities at Robert Crown Memorial Beach are kite surfing and wind surfing at high tide and wading, exploring, and playing in shallow pools at low tide. In part because of cool water temperatures at this beach, swimming is uncommon, and kite and wind surfers usually wear wetsuits. Most dermatitis cases at this beach were contracted by persons wading in tide pools. Dermatitis usually occurred on the feet or legs. Only 1 case was reported among kite surfers and wind surfers. Since 2005, three biologists working at this beach have contracted dermatitis, usually on their hands or forearms (Figure 6). In experiments with G. huttoni schistosomes obtained near Miami, Florida, cercariae emerged only when temperatures were >22°C, regardless of season or light intensity (32). In San Francisco Bay, the highest numbers of cercariae from H. japonica may be released in beach flat tide pools that warm up during daytime low tides.
There are at least 3 ways in which schistosomes we found in H. japonica snails could have recently emerged as a disease agent in San Francisco Bay. First, an unknown native schistosome may be present in native birds and snails, which switched hosts to the introduced H. japonica snail. Some evidence shows that schistosomes are capable of switching hosts (39). Neither of 2 native Haminoea species found in western North America between Baja, California and Alaska (H. vesicula [Gould 1855] and H. virescens) (22) is known to host schistosomes or is found in the study area. However, if an undetected native schistosome is present in 1 of these species, or in a Haminoea species further south on the Pacific Coast, migration of its bird host over San Francisco Bay could have resulted in infection of the H. japonica population at Robert Crown Memorial Beach, greatly increasing the potential for human dermatitis. The range of G. huttoni schistosomes may extend from Florida through the Caribbean and (by bird movements) up the Pacific Coast.
Second, the schistosomes could be a species from Asia recently introduced into San Francisco Bay in infected H. japonica snails. This introduction would require adult snails (eggs and larvae do not contain schistosomes) that harbor male and female cercariae; both sexes would have to colonize the same bird to initiate egg-producing infections. This event occurred on 1 occasion when A. variglandis schistosomes and I. obsoleta snails were introduced into western North America. Whether H. japonica snails arrived in San Francisco Bay directly from Asia or indirectly through Washington State is unknown. H. japonica snails may have been introduced into Washington State, where they were first observed in the early 1980s (40), in Pacific oysters (Crassostrea gigas Thunberg 1793) imported from Japan for mariculture (17). Pacific oysters from hatcheries or oyster farms in Japan, Washington, Oregon, and California were placed in San Francisco Bay for commercial mariculture in the 1930s, for occasional experimental use until 1981, for bioaccumulation studies during 1991–2002, and were introduced illegally at 1 site in 1999. A population recently discovered in South San Francisco Bay appears to have been introduced during the late 1990s (A.N. Cohen, D. Goodwin, unpub. data). These occurrences may be related to the initial appearance of H. japonica snails in 1999 in South San Francisco Bay.
Third, the schistosomes could be a species from Asia found in migrating birds that infected H. japonica snails after these snails became established. Because some birds excrete schistosome eggs for <10–28 months postinfection (19,23), some schistosomes may survive in a bird host long enough to complete a long-distance migration. However, because no bird species are known to routinely migrate across the Pacific Ocean between the native region of H. japonica snails in Asia and regions in the western United States, introduction by this mechanism seems unlikely.
Much remains to be learned about factors favoring outbreaks of cercarial dermatitis in new areas. Native Haminoea spp. should be surveyed for parasites to assess whether host switching may be involved, and H. japonica snails should be surveyed in their native range and in Washington State to determine whether trans-Pacific schistosome colonization events have occurred and by what mechanisms. Molecular analysis of G. huttoni schistosomes would increase the taxonomic status of the species we isolated from San Francisco Bay. The definitive avian host in this region could be determined by examination of feces for eggs and carcasses for adult schistosomes.
The molecular signatures we have provided may be present in schistosomes isolated from birds or snails in other areas, which would help establish how this zoonotic infection reached California. Potential effects on native biota, especially endangered birds that might serve as hosts (such as the California least tern or California brown pelican), should be assessed. Whether this schistosome will become established in other locations along the Pacific Coast and affect beach users is unknown. Improved understanding of the biology and mechanism of establishment of this schistosome may enable better management of human exposure and infection, control of its spread, and prevention of other schistosome introductions or outbreaks.
Dr Brant is a research assistant professor at the University of New Mexico. Her main research interests are epidemiology of cercarial dermatitis and evolutionary history of avian schistosomes.
Acknowledgments
We thank Steve Stricker for collecting H. virescens snails, Jitka Aldhoun for help with dissecting and examining snails, Zachary Phillips for help with examining parakeets, Dave Hammett for collecting seagulls, and the San Francisco Estuary Institute for providing laboratory space.
This study was supported by the College of Arts and Sciences at the University of New Mexico through National Institutes of Health (NIH) grants RO1 AI44913 and P20 RR018754. Technical assistance at the University of New Mexico Molecular Biology Facility was supported by NIH grant 1P20RR18754 from the Institute Development Award Program of the National Center for Research Resources. The Swimmer Itch Hotline was supported by the California State Beach Water Monitoring Fund.
References
- Grodhaus G, Keh B. The marine dermatitis-producing cercaria of Austrobilharzia variglandis in California (Trematoda: Schistosomatidae). J Parasitol. 1958;44:633–8. DOIPubMedGoogle Scholar
- Lafferty KD, Smith KF, Torchin ME, Dobson AP, Kuris AM. The role of infectious disease in natural communities: what introduced species tell us. In: Sax DF, Stachowicz JJ, Gaines SD, editors. Species invasions: insights into ecology, evolution, and biogeography. Sunderland (MA): Sinauer Associates; 2005. p. 111–34.
- Sauer JS, Cole RA, Nissen JM. Finding the exotic faucet snail (Bithynia tentaculata): investigation of waterbird die-offs on the Upper Mississippi River National Wildlife and Fish Refuge: U.S. Geological Survey Open-File Report 2007–1065. Washington: The Survey; 2007.
- Miura O, Torchin ME, Kuris AM, Hechinger RF, Chiba S. Introduced cryptic species of parasites exhibit different invasion pathways. Proc Natl Acad Sci U S A. 2006;103:19818–23. DOIPubMedGoogle Scholar
- Torchin ME, Byers JE, Huspeni TC. Differential parasitism of native and introduced snails: replacement of a parasite fauna. Biological Investigations. 2005;7:885–94. DOIGoogle Scholar
- Telfer S, Bown KJ, Sekules R, Begon I, Hayden T, Birtles R. Disruption of a host-parasite system following the introduction of an exotic host species. Parasitology. 2005;130:661–8. DOIPubMedGoogle Scholar
- Wyatt KB, Campos PF, Gilbert M, Thomas P, Gilbert P, Kolokotronis SO, Historical mammal extinction on Christmas Island (Indian Ocean) correlates with introduced infectious disease. PLoS ONE. 2008;3:e3602. DOIPubMedGoogle Scholar
- Torchin ME, Lafferty KD, Dobson AP, McKenzie VJ, Kuris AM. Introduced species and their missing parasites. Nature. 2003;421:628–30. DOIPubMedGoogle Scholar
- Friend M, McLean RG, Dein FJ. Disease emergence in birds: challenges for the twenty-first century. Auk. 2001;118:290–303. DOIGoogle Scholar
- Carlton JT. San Francisco Bay—the urbanized estuary: investigations into the natural history of San Francisco Bay and Delta with reference to the influence of man. Conomos TJ, editor. Symposium held in conjunction with the Annual Meeting of the Pacific Section of The American Society of Limnology and Oceanography, June 12–16, 1977, in affiliation with the 58th Annual Meeting of the Pacific Division, American Association for the Advancement of Science, San Francisco, Pacific Division. Washington: American Association for the Advancement of Science; 1979. p. 427–44.
- Cohen AN, Carlton JT. Biological study. Nonindigenous aquatic species in a United States estuary: a case study of the biological invasions of the San Francisco Bay and Delta. NTIS PB96–166525. Washington: US Fish and Wildlife Service and the National Sea Grant College Program, Connecticut Sea Grant; 1995.
- Cohen AN, Carlton JT. Accelerating invasion rate in a highly invaded estuary. Science. 1998;279:555–8. DOIPubMedGoogle Scholar
- Penner LR. Cercaria littorinalinae sp. nov., a dermatitis-producing schistosome larva from the marine snail, Littorina planaxis Philippi. J Parasitol. 1950;36:466–72. DOIPubMedGoogle Scholar
- Stunkard HW, Hinchliffe MC. The morphology and life history of Microbilharzia variglandis (Miller and Northup, 1926) Stunkard and Hinchliffe, 1951, avian blood flukes whose larvae cause “swimmer’s itch” of ocean beaches. J Parasitol. 1952;38:248–65. DOIPubMedGoogle Scholar
- Arnold HL, Bonnet DD. Swimmer’s itch, its first appearance in Hawaii. Proceedings of the Hawaiian Academy of Science. 1950;25:4.
- Penner LR. The biology of a marine dermatitis producing schistosome cercaria from Batillaria minima. J Parasitol. 1953;39:19–20.
- Rohde K. The bird schistosome Austrobilharzia terrigalensis from the Great Barrier Reef, Australia. Z Parasitenkd. 1977;52:39–51. DOIPubMedGoogle Scholar
- Tang Z, Tang C. Dermatitis producing schistosomes of birds and mammals in China [in Chinese]. Acta Zoologica Sinica. 1976;22:341–60.
- Appleton CC. Studies on Austrobilharzia terrigalensis (Trematoda: Schistosomatidae) in the Swan Estuary, Western Australia: infection in the definitive host Larus novaehollandiae. Int J Parasitol. 1983;13:249–59. DOIPubMedGoogle Scholar
- Leighton BJ, Ratzlaff D, McDougall C, Stewart G, Nadan A, Gustafson L. Schistosome dermatitis at Crescent Beach, preliminary report. Environmental Health Review. 2004;48:5–13.
- Bearup AJ. A schistosome larva from the marine snail Pyrazus australis as a cause of cercarial dermatitis in man. Med J Aust. 1955;42:955–60.PubMedGoogle Scholar
- Gosliner TM, Behrens DW. Anatomy of an invasion: systematics and distribution of the introduced opisthobranch snail, Haminoea japonica Pilsbry, 1895 (Gastropoda: Opisthobranchia: Haminoeidae). Proc Calif Acad Sci. 2006;57:1003–10.
- Leigh WH. The morphology of Gigantobilharzia huttoni (Leigh, 1953), an avian schistosome with marine dermatitis producing larvae. J Parasitol. 1955;41:262–9. DOIPubMedGoogle Scholar
- Brant SV, Morgan JAT, Mkoji GM, Snyder SD, Rajapakse JR, Loker ES. An approach to revealing blood fluke life cycles, taxonomy, and diversity: provision of key reference data including DNA sequence from single life cycle stages. J Parasitol. 2006;92:77–88. DOIPubMedGoogle Scholar
- Brant SV, Loker ES. Molecular systematics of the avian schistosome genus Trichobilharzia (Trematoda: Schistosomatidae) in North America. J Parasitol. 2009;95:941–63. DOIPubMedGoogle Scholar
- Swofford DL. PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods). Sunderland (MA): Sinauer Associates; 2002.
- Ronquist F, Huelsenbeck JP. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–4. DOIPubMedGoogle Scholar
- Posada D. jModelTest: phylogenetic model averaging. Mol Biol Evol. 2008;25:1253–6. DOIPubMedGoogle Scholar
- Leigh WH. Cercaria huttoni, sp. nov., a dermatitis-producing schistosome larva from the marine snail, Haminoea antillarum guadalupensis Sowerby. J Parasitol. 1953;39:625–9. DOIPubMedGoogle Scholar
- Najim AT. Life history of Gigantobilharzia huronensis Najim, 1950, a dermatitis-producing bird blood-fluke (Trematoda-Schistosomatidae). Parasitology. 1956;46:443–69. DOIPubMedGoogle Scholar
- Miller HM Jr, Northup FE. The seasonal infestation of Nassa obsoleta (Say) with larval trematodes. Biol Bull. 1926;50:490–509. DOIGoogle Scholar
- Hutton RF. Schistosome cercariae as the probable cause of seabather’s eruption. Bull Mar Sci Gulf Caribb. 1952;2:346–59.
- Ewers WH. A new intermediate host of schistosome trematodes from New South Wales. Nature. 1961;190:283–4. DOIPubMedGoogle Scholar
- Martorelli SR, Alda P. Larval digeneans of the siphonariid pulmonates Siphonaria lessoni and Kerguelenella lateralis and the flabelliferan isopod Exosphaeroma sp. from the intertidal zone of the Argentinean Sea. Comp Parasitol. 2009;76:267–72. DOIGoogle Scholar
- Khalifa R. Studies on schistosomatidae Looss. (Trematoda) of aquatic birds of Poland II. Gigantobilharzia mazuriana sp. n., with a discussion of the subfamily Gigantobilharziinae Mehra, 1940. Acta Parasitologica Polonica. 1899;1974:265–84.
- Leigh WH. Brown and white pelicans as hosts for schistosomes of the genus Gigantobilharzia. J Parasitol. 1957;43:35–6.
- Kinsella JM, Spalding MG, Forrester DJ. Parasitic helminths of the American White Pelican, Pelecanus erythrorhynchos, from Florida, U.S.A. Comp Parasitol. 2004;71:29–36. DOIGoogle Scholar
- Cort WW. Studies on schistosome dermatitis XI. Status of knowledge after more than twenty years. Am J Hyg. 1950;52:251–307.PubMedGoogle Scholar
- Brant SV, Loker ES. Can specialized pathogens colonize distantly related hosts? Schistosome evolution as a case study. PLoS Pathog. 2005;1:167–9. DOIPubMedGoogle Scholar
- Gibson GD, Chia F. Description of a new species of Haminoea, Haminoea callidegenita (Mollusca: Opisthobranchia), with a comparison with two other Haminoea species found in the northeast Pacific. Can J Zool. 1989;67:914–22. DOIGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 16, Number 9—September 2010
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Sara V. Brant, Department of Biology, University of New Mexico, 1 University of New Mexico, MSC03 2020, Albuquerque, NM, 87131, USA
Top