Volume 19, Number 12—December 2013
Research
Antiviral Susceptibility of Highly Pathogenic Avian Influenza A(H5N1) Viruses Isolated from Poultry, Vietnam, 2009–2011
Cite This Article
Citation for Media
Abstract
We assessed drug susceptibilities of 125 avian influenza A(H5N1) viruses isolated from poultry in Vietnam during 2009–2011. Of 25 clade 1.1 viruses, all possessed a marker of resistance to M2 blockers amantadine and rimantadine; 24 were inhibited by neuraminidase inhibitors. One clade 1.1 virus contained the R430W neuraminidase gene and reduced inhibition by oseltamivir, zanamivir, and laninamivir 12-, 73-, and 29-fold, respectively. Three of 30 clade 2.3.4 viruses contained a I223T mutation and showed 7-fold reduced inhibition by oseltamivir. One of 70 clade 2.3.2.1 viruses had the H275Y marker of oseltamivir resistance and exhibited highly reduced inhibition by oseltamivir and peramivir; antiviral agents DAS181 and favipiravir inhibited H275Y mutant virus replication in MDCK-SIAT1 cells. Replicative fitness of the H275Y mutant virus was comparable to that of wildtype virus. These findings highlight the role of drug susceptibility monitoring of H5N1 subtype viruses circulating among birds to inform antiviral stockpiling decisions for pandemic preparedness.
Sporadic transmission of highly pathogenic avian influenza (HPAI) A(H5N1) viruses from birds to humans has been documented since 1997 (1), and these viruses continue to cause severe illness and death in humans. Their wide geographic spread and rapid evolution have raised concerns over emergence of a novel, virulent virus that could efficiently transmit among humans, leading to a pandemic. Vietnam is among the countries experiencing the highest number of human fatalities caused by zoonotic H5N1 subtype infections. Since the introduction of HPAI (H5N1) viruses into poultry in Vietnam during 2003 (1,2), there have been dynamic changes in their genetic and antigenic properties. Clade 1 viruses predominated in Vietnam before 2007, and were the most commonly detected H5N1 subtype group in the Mekong Delta region through 2010 (3). However, in northern Vietnam provinces, clade 2.3.4 viruses became the predominant group during 2007–2010. Since 2010, viruses of clade 2.3.2.1 have been detected in poultry from both regions (3). Since 2009, multiple subgroups of 2.3.2.1 rapidly emerged and have circulated among domestic poultry in Asia, including several provinces of Vietnam (4).
Genetic and antigenic divergence of HPAI (H5N1) viruses among poultry challenges development of effective vaccines for poultry and to pandemic preparedness and development of antiviral drugs for humans. Assessment of drug susceptibility has become an integral part of subtype H5N1 virus surveillance. To assist laboratories worldwide in their surveillance and pandemic preparedness efforts, the Influenza Division of the Centers for Disease Control and Prevention (CDC), Atlanta, GA, USA, along with other partners, developed the H5N1 Genetic Changes Inventory that includes established and potential markers of drug resistance (5). Resistance to matrix 2 (M2) protein blockers amantadine and rimantadine, caused by mutations in the M2 protein, is detected commonly in clade 1.1 (S31N) and clade 2.1.3 (V27A) H5N1 virus subtypes and sporadically in other groups (6,7). Oseltamivir, an orally administered neuraminidase (NA) inhibitor, is the most prescribed medication for the treatment of persons with influenza virus infections. Emergence of resistance to NA inhibitors among H5N1 virus subtypes, especially oseltamivir resistance among H5N1 subtypes caused by the H275Y mutation, is a constant threat (8). Assessment of susceptibility to NA inhibitors is hampered by several factors: insufficient knowledge of molecular markers of resistance, lack of harmonized approaches for testing and data analysis and, most critically, lack of established laboratory correlates of clinically relevant resistance. Taking into account these and other limitations, the current method for monitoring susceptibility to NA inhibitors is a critical element needed to evaluate pandemic risk.
In this study, we assessed drug susceptibility profiles of HPAI A(H5N1) viruses isolated from poultry specimens collected in Vietnam during 2009–2011. The antiviral drugs tested included FDA-approved medications and investigational antiviral agents. We report the detection of an oseltamivir-resistant virus with H275Y mutation from the expanding clade 2.3.2.1.
Viruses
Viruses collected from poultry on farms, in backyard flocks, and in live-poultry markets in Vietnam during 2009–2011 were identified as HPAI A(H5N1) at the National Center for Veterinary Diagnostics (NCVD), Vietnam, by using the World Health Organization (WHO) protocol (9). Viruses were then sent to the WHO Collaborating Center for Surveillance, Epidemiology and Control of Influenza at CDC, where they were isolated from eggs and further propagated according to WHO protocol (9). Virus handling was conducted under enhanced Biosafety Level 3 containment according to institutional guidelines.
Sequencing and Phylogenetic Analysis
Full-length gene sequences were generated by the ABI PRISM 3730 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) and assembled by using Sequencher 5.0 (Gene Codes Corporation, Ann Arbor, MI, USA). Phylogenetic trees were generated by using MEGA version 5.0 (www.megasoftware.net) neighbor-joining methods implemented with 1,000 bootstrap replicates. Phylogenetic data for the strain A/goose/Guangdong/1/1996 (clade 0) were used as a reference for tree rooting and numbering, and trees were annotated according to the WHO/World Organisation for Animal Health/Food and Agriculture Organization of the United Nations criteria (10). Sequences were deposited into the Global Initiative on Sharing All Influenza Data database. Accession numbers are listed in Technical Appendix 1. For NA sequences, N1 aa numbering is used throughout the text (11). The pyrosequencing method was used to detect NA residue 275 in inoculated ferret nasal wash samples (12).
NA Inhibitors, Neuraminidase Inhibition Assay, and 50% Inhibitory Concentration Analysis
Susceptibility to the drugs zanamivir (GlaxoSmithKline, Uxbridge, UK), oseltamivir (Roche Diagnostics GmbH, Mannheim, Germany), peramivir (BioCryst Pharmaceuticals, Birmingham, AL, USA), and laninamivir (compound R-125489; Biota, Begbroke, UK) was assessed by fluorescent neuraminidase inhibition (NI) assay, by using inhibitor concentrations ranging from 0.03 nmol/L to 1,000 nmol/L (13). The 50% inhibitory concentration (IC50) values, the drug concentration needed to inhibit virus NA activity by 50%, were determined by using a CDC in-house program, the JASPR v1.2 curve-fitting software (14). Statistical analysis of IC50 values was performed by using SAS 9.2 software (SAS Institute, Cary, NC, USA) to identify outliers, using a statistical cutoff value U = Q3+3.0*(interquartile range). The interquartile range was determined as Q3–Q1; Q1 or Q3 denoted 25th or 75th percentile, respectively. The resulting value was applied for clade and drug. Mild outliers were defined as viruses that had IC50 values >U and <10 times the median, and extreme outliers as having IC50values >U and ≥10 times the median. SigmaPlot 12 (Systat Software, Chicago, IL, USA) was used to generate box-and-whisker plots to visualize outliers. The median/mean IC50 values among virus clades were analyzed by using the Kruskal-Wallis 1-way analysis of variance and the Dunn’s multiple comparison test, respectively.
Susceptibility to Antiviral Agents in Cell Culture
Susceptibilities to the M2 blocker amantadine and to investigational agents DAS181 and favipiravir (T705) were assessed in a virus yield reduction assay on Madin-Darby canine kidney (MDCK) SIAT1 cells (15,16). In brief, the confluent cell monolayers seeded on 96-well plates were treated before inoculation with amantadine or favipiravir for 30 minutes or with DAS181 for 2 hours. After the drug was removed, cells were inoculated with either wildtype (WT) virus, which did not contain the H275Y mutation in NA, or oseltamivir-resistant virus with the H275Y mutation (as positive control) at a low multiplicity of infection of 0.0001 PFU/cell and incubated for 1 hour at 4°C. Cells were washed and added to fresh media containing drug dilutions, and incubated at 37°C for 24 hours. Supernatants were harvested to determine infectious virus yield 50% tissue culture infectious dose per mL (TCID50/mL) in MDCK-SIAT1 cells. The 90% effective concentration (EC90) of drug (drug concentration that reduces the infectious virus yield by 90%) was determined by using the 4-parameter logistic nonlinear regression model equation in GraphPad Prism 5 software (GraphPad Software, La Jolla, CA, USA).
Replicative Capacity in Cell Culture and Ferret Upper Respiratory Tract
Comparison of replicative capacity of WT and oseltamivir-resistant H275Y mutant virus was achieved by inoculating MDCK-SIAT1 cells at a multiplicity of infection of 0.0001 PFU/cell in 24-well plates. Cells were incubated at 37°C, and supernatants were collected every 12 hours until 72 hours postinoculation. At each time point, the infectious virus yields (TCID50/mL) were determined by titrating the supernatants on MDCK-SIAT1 cells (17). In vivo replicative capacity was assessed in ferrets by inoculating naïve, anesthetized animals, 3–5 months of age, with 106 TCID50 of either WT or H275Y virus. Virus titers (TCID50/mL) were measured in the nasal washes collected postinoculation on days 1–3, 5, 7, and 9.
Phylogenetic Analysis and Resistance Markers
Figure 1

Figure 1. . Description of 125 highly pathogenic avian influenza A(H5N1) viruses collected from poultry in Vietnam during 2009–2011 and tested during this study.
Figure 2

Figure 2. . Distribution of log-transformed 50% inhibitory concentration (IC50) values for oseltamivir, zanamivir, and peramivir: Box-and-whisker plot analysis of all tested highly pathogenic avian influenza A(H5N1) viruses (n = 125) and individual...
Knowledge of subtype H5N1 virus genomics, especially that of hemagglutinin (HA), NA, and membrane protein, or M genes, is required for interpretation of drug susceptibility data and for uncovering new trends. According to the HA gene phylogeny, viruses isolated from poultry in Vietnam during 2009–2011 were assigned to 3 clades: 1.1 (n = 25), 2.3.2.1 (n = 70), and 2.3.4 (n = 30) (Figure 1; Technical Appendix 2, Figure 2, panel A. The viruses of clade 2.3.4 were categorized as 3 subclades, termed 2.3.4.1, 2.3.4.2, and 2.3.4.3. The NA phylogenetic tree topology (Technical Appendix 2, Figure 2, panel B) was comparable to the HA tree for clades 1.1 and 2.3.2.1, but not for clade 2.3.4 viruses, suggesting NA gene reassortment among these viruses. Two reassortant viruses of subclade 2.3.4.1 contained NA genes similar to those of subclade 2.3.2.1 viruses.
The M tree was similar to the HA tree and showed evidence of reassortment for a single clade 2.3.4.2 virus that clustered with M genes from subclade 2.3.4.1 (Technical Appendix 2, Figure 2, panel C). The H5N1 Genetic Changes Inventory (5) was used to screen NA and M gene alignments for molecular markers associated with potential drug resistance. Clade 1.1 viruses contained S31N in M2 protein, the most common marker of M2 blocker resistance. This mutation was present in combination with L26I, a typical feature of clade 1 and 1.1 viruses. The remaining viruses had no known markers of M2 blocker resistance. In the NA gene, the H275Y mutation, a marker of oseltamivir-resistance, was detected in the virus A/duck/VN/NCVD-664/2010 (clade 2.3.2.1). The virus was isolated in Ninh Binh Province in northern Vietnam. In subclade 2.3.4.2 (n = 13), 3 viruses contained the mutation I223T, which may affect susceptibility to NA inhibitors. One clade 1.1 virus carried the mutation V149A, which was previously linked to slightly reduced susceptibility to zanamivir (18).
Drug Susceptibility in NI Assay
To identify viruses with potential resistance to NA inhibitor(s), we performed the NI assay using oseltamivir, zanamivir, and peramivir. IC50 values were calculated for each virus and drug. The median IC50 values were similar for oseltamivir and zanamivir (0.44 nmol/L and 0.36 nmol/L, respectively) and ≈2-fold lower for peramivir (Technical Appendix 2, Table). The influence of the NA sequence diversity on IC50 values was most noticeable from the wide-ranging oseltamivir IC50 values (>13,000-fold difference between minimum and maximum), whereas the range was much narrower for zanamivir (145-fold) and intermediate for peramivir (1,300-fold). Analysis of IC50 values was further achieved by individual clade to enable better correlation with NA sequences (Table 1; Figure 2). The 2 reassortants were excluded from clade 2.3.4 analysis because the associated NA gene from each was related to those from clade 2.3.2.1.
The median oseltamivir IC50 value for clade 1.1 was lower than that of clades 2.3.2.1 and 2.3.4, by 7- to 27-fold, respectively. In clade 1.1, virus A/chicken/Vietnam/NCVD-780/2011 was identified as an extreme outlier and showed a 12-fold increased oseltamivir IC50 value (Table 2; Figure 2). This virus contained a previously unreported change: the presence of NA mutation R430W. Virus A/chicken/Vietnam/NCVD-776/2011 was identified as a mild outlier: it had the H253Y mutation and showed a 9-fold increase in oseltamivir IC50 values; virus A/chicken/Vietnam/NCVD-878/2011, which carried V149A, exhibited a 3-fold increase. Notably, the oseltamivir IC50 values of all 3 outliers described were similar to, or less than, the median oseltamivir IC50 value of clade 2 viruses (Tables 1, 2; Figure 2).
Influenza virus strain H5N1 A/duck/Vietnam/NCVD-664/2010 was identified as an extreme outlier for oseltamivir susceptibility in clade 2.3.2.1; it contained the marker H275Y and exhibited a 1,353-fold elevation in IC50. Two mild outliers (3–5-fold increase) that carried the V424I change were identified within the same clade. In clade 2.3.4 viruses, 4 outliers for oseltamivir were detected, 3 of which possessed I223T, which conferred a 6–7-fold increase in IC50 values. The fourth virus had a V147R substitution and exhibited a 4-fold increase in IC50 (Table 2). As anticipated from the results of phylogenetic analysis, oseltamivir IC50 values of the 2 reassortant viruses (HA of clade 2.3.4 but NA from clade 2.3.2.1) matched those of clade 2.3.2.1 viruses (Table 1).
When tested for zanamivir susceptibility, an extreme outlier that had a 73-fold increase in IC50 was detected in clade 1.1 (Table 2): this was the same virus, A/chicken/Vietnam/NCVD-780/2011, that showed a previously unknown R430W change and was identified as an extreme outlier for oseltamivir susceptibility. Three mild outliers were identified from clades 1.1, 2.3.2.1, and 2.3.4 and had amino acid changes at the V149A, H275Y, and G147R substitutions, respectively.
The virus A/duck/Vietnam/NCVD-664/2010 that carried the H275Y mutation was predictably identified as an extreme outlier for peramivir with a 415-fold increase in IC50 values; the remaining viruses showed no increase. Among a subset of viruses (n = 38) tested with laninamivir, the virus that carried the R430W mutation showed a 29-fold increase, and the virus that had the H275Y mutation showed a 6-fold increase in IC50 values.
The WHO criteria for reporting NI assay data for influenza viruses (19) are based on fold difference between IC50 values of the test virus and a reference IC50 value (such as median IC50); different criteria are set for seasonal type A and type B viruses. The reporting for H5N1subtypes is not specified; therefore, we followed the criteria as outlined for seasonal type A viruses, but grouped the IC50 values by clade (Table 1). For clade 1.1, the virus that had the R430W mutation showed reduced inhibition by oseltamivir, zanamivir, and laninamivir; in clade 2.3.2.1, the virus that had the H275Y mutation showed highly reduced inhibition by oseltamivir and peramivir.
Characterization of the Oseltamivir-Resistant H275Y Virus
Figure 3
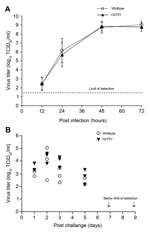
Figure 3. . Replicative capacity of the oseltamivir-resistant highly pathogenic avian influenza A(H5N1) virus possessing the H275Y substitution and the wild type virus in (A) MDCK and MDCK-SIAT1 cell lines and (B) in...
The oseltamivir-resistant virus was also tested with antiviral agents with mechanisms of action other than NA inhibition. The infectious virus yields of WT and the oseltamivir-resistant virus were reduced by >2 logs at 1 µg/mL of amantadine (data not shown), which is consistent with the M2 blocker–sensitive genotype. Inoculation of cells with DAS181 before incubation was equally effective in inhibiting replication of the virus with H275Y mutation and the WT virus (Table 3). Both viruses were equally susceptible to favipiravir, expressing EC90 values of 3 µmol/L–6 µmol/L (Table 4). For risk assessment, it was essential to investigate whether the H275Y mutation had a detrimental effect on virus replication. In MDCK-SIAT1 cells, the H275Y-mutated virus replicated at a similar rate as the WT virus and reached infectious titers as high as 109 TCID50/mL at 72 hours post infection (Figure 3, panel A). In the ferret model, the H275Y virus titers in the nasal washes collected at several points after inoculation were similar to those of the WT virus (Figure 3, panel B) and no differences in symptoms were noted. The stability of the H275Y mutation was demonstrated by analyzing pyrosequencing data of the viruses shed by the infected ferrets.
In this study, we identified a single oseltamivir-resistant virus among 125 HPAI A(H5N1) viruses isolated from poultry in Vietnam during 2009–2011. It was recovered from a domestic duck in Ninh Binh Province, northern Vietnam. This virus belonged to the rapidly expanding clade 2.3.2.1 and contained the H275Y mutation, which is the principal marker of oseltamivir resistance in N1-subtype viruses (11). We observed no impairment in its replicative fitness in either cell culture or a ferret model. Our findings are in agreement with previous studies in which the reverse genetically engineered H275Y mutant from clade 1 retained its in vitro replicative efficiency and high pathogenicity in animals (20). Emergence of clade 1 viruses carrying the H275Y mutation was reported in oseltamivir-treated patients (21,22). When tested in ferrets, replication of the H275Y mutant virus was not inhibited by oseltamivir (23), confirming its oseltamivir-resistant phenotype. Of the 3,215 subtype H5N1 virus NA sequences available in GenBank, 6 contain H275Y; these viruses include a virus isolated from a patient treated with oseltamivir in Vietnam during 2005 (21), a patient in Indonesia (GenBank accession no. EU146786; unknown treatment history), and 4 viruses isolated from birds in Hong Kong (GenBank accession no. DQ250158) and Russia (GenBank accession no. DQ840522, DQ320136, and CY063862) (7). Although detection of H275Y mutations in H5N1 subtypes is rare, the global spread of subtype H1N1 viruses carrying the same mutation in the absence of drug exposure serves as a sobering reminder of the unpredictable nature of influenza virus evolution. As indicated in this and other studies, the HPAI (H5N1) NA gene is subject to reassortment between different HA clade-bearing viruses, which could accelerate this process (3).
Apart from the H275Y marker, it is difficult to predict the effect of natural genetic variation in HPAI (H5N1) viruses on susceptibility to NA inhibitors in humans. The range of H5N1 subtype oseltamivir IC50 values was wide (0.04–527.26 nmol/L), similar to previous findings (24). The WHO criteria for reporting NI assay data are based on a fold increase in the IC50 value of a test virus compared with that of a control or reference virus (or median) IC50 value (18). For influenza A, the result is interpreted as normal (<10-fold), reduced (10- to100-fold), or highly reduced inhibition (>100-fold).
In this report, the subtype H5N1 influenza virus with the H275Y NA mutation exhibited highly reduced inhibition by oseltamivir and peramivir, and normal inhibition by zanamivir and laninamivir. The current criteria need further clarification when applied to highly diverse H5N1 subtypes. For instance, oseltamivir IC50 values of clade 1.1 viruses were 7–27-fold lower than those of clade 2 viruses; this finding is in accord with previous reports (25). This difference stems from the presence of either histidine (clade 2) or tyrosine (clade 1) at position 253 in the NA protein (26,27). Predictably, the identified clade 1.1 virus carrying the revertant H253Y mutation exhibited ≈9-fold higher IC50 values compared with the median IC50 value for this clade and ≈10-fold increase compared with A/ck/VN/NCVD-777/2011, a matching virus in which the only difference in the NA gene was an H253Y substitution. Such a virus would be reported as exhibiting reduced inhibition, yet its oseltamivir IC50 value was below the median IC50 value of typical clade 2.3.4 viruses tested in this study. In view of these findings, it may be more appropriate to consider all clade 1 viruses possessing histidine at position 253 as hypersensitive and use a median IC50 value of clade 2 viruses in fold difference calculations. If this rule were applied, 1 of the reference H5N1 subtype viruses (clade 1), possessing N295S (23), would increase by ≈2-fold in oseltamivir IC50 values (2.99 nmol/L) compared with values of clade 2.3.4 viruses (1.62 nmol/L) and, thus, would be reported as exhibiting normal inhibition. Like the H275Y-mutated viruses previously identified, this variant has also been isolated from an oseltamivir-treated patient (22) and its replication was not affected by oseltamivir treatment in ferrets (23). Therefore, it is essential to be able to detect and report these NA variants (28), despite an overall low IC50 value.
Because of lack of established laboratory correlates of clinically relevant resistance, analysis and interpretation of IC50 values generated in this study were completed according to the WHO criteria with a stipulation that fold change comparison was performed by using median IC50 values for individual clades. This approach facillitated identification of variants, such as the R430W-mutated virus from clade 1.1, which should be further studied. Certain discrepancies among reports are to be expected in the absence of standardized assays and criteria. For example, variant V149A of clade 1.1 was reported here as normally inhibited by zanamivir; such a variant was previously reported as showing mildly decreased susceptibility to zanamivir (18,29). Similarly, substitutions at variant I223 have previously been associated with reduced susceptibility to oseltamivir, zanamivir, or both (5). In this study, 3 clade 2.3.4 viruses carrying variant I223T were reported to show normal inhibition by NA inhibitors because of the relatively high median IC50 values of this clade. Nevertheless, the oseltamivir IC50 values in this clade were ≈55-fold greater than the reference WT A(H1N1)pdm09 virus. Acquired additional changes in the NA (e.g., H275Y) may confer a higher level of resistance to the NA-inhibitor class of drugs among these viruses (30,31).
HPAI (H5N1) viruses resistant to M2 blockers are prevalent among poultry throughout Asia, including Vietnam (7,32). Most of these M2-resistant viruses belong to clade 1.1 (32,33), but they have also been found in other clades, including clade 2.3.4 (34). The oseltamivir-resistant H275Y-mutant virus detected in this study was sensitive to M2 blockers; however, the ease with which viruses can acquire resistance to this class of drugs emphasizes the need for alternative therapeutic options. The NA activity of the oseltamivir-resistant H275Y-mutant virus was inhibited by zanamivir and laninamivir in the NI assay. These NA inhibitors are delivered by inhalation, which limits their use for treatment of severely ill patients; an intravenous formulation of zanamivir is in clinical trial and is available on a compassionate use basis for treatment of hospitalized influenza patients (35,36). The replication of the H275Y-mutant virus and its WT counterpart were equally inhibited by investigational drugs DAS181 and favipiravir in cell culture.
Our findings demonstrate the critical role of ongoing monitoring of antiviral drug susceptibility in HPAI (H5N1) viruses sampled from poultry on informing antiviral stockpiling decisions for pandemic preparedness. Because 15 countries have reported human cases of HPAI (H5N1) virus infection to date, these findings also emphasize the need to enhance the armamentarium of available anti-influenza drugs worldwide for treatment of subtype H5N1-infected patients, including agents with diverse mechanisms of action, which could enable combination treatment (37), and host-directed antiviral therapy, and which may be less vulnerable to resistance.
Dr Ha T. Nguyen is a Battelle research scientist on the Molecular Epidemiology Team of the Influenza Division, Centers for Disease Control and Prevention, in Atlanta, Georgia. Her research interests include influenza viruses and mechanisms of resistance to antiviral agents.
Acknowledgment
We thank our collaborators in the National Center for Veterinary Diagnostics, Hanoi, Vietnam, for their valuable contributions to this study. We also thank Ronald B. Moss and Yousuke Furuta for kindly providing investigational anti-influenza drugs DAS181 and favipiravir, respectively, and Anton Chesnokov for his excellent technical assistance.
References
- Wan XF, Nguyen T, Davis CT, Smith CB, Zhao ZM, Carrel M, Evolution of highly pathogenic H5N1 avian influenza viruses in Vietnam between 2001 and 2007. PLoS ONE. 2008;3:e3462. DOIPubMedGoogle Scholar
- Dung Nguyen T, Nguyen TV, Vijaykrishna D, Webster RG, Guan Y, Malik Peiris JS, Multiple sublineages of influenza A virus (H5N1), Vietnam, 2005–2007. Emerg Infect Dis. 2008;14:632–6. DOIPubMedGoogle Scholar
- Nguyen T, Rivailler P, Davis CT. Hoa do T, Balish A, Dang NH, et al. Evolution of highly pathogenic avian influenza (H5N1) virus populations in Vietnam between 2007 and 2010. Virology. 2012;432:405–16.
- World Health Organization. Antigenic and genetic characteristics of zoonotic influenza viruses and development of candidate vaccine viruses for pandemic preparedness. Influenza, Vaccines, Vaccine Viruses. February 2013 [cited 2013Aug 6]. http://www.who.int/influenza/vaccines/virus/characteristics_virus_vaccines/en/
- Centers for Disease Control and Prevention. H5N1 Genetic changes inventory. [cited 2013 Aug 6]. http://www.cdc.gov/flu/avianflu/h5n1/inventory.htm
- Cheung CL, Rayner JM, Smith GJ, Wang P, Naipospos TS, Zhang J, Distribution of amantadine-resistant H5N1 avian influenza variants in Asia. J Infect Dis. 2006;193:1626–9. DOIPubMedGoogle Scholar
- Hill AW, Guralnick RP, Wilson MJ, Habib F, Janies D. Evolution of drug resistance in multiple distinct lineages of H5N1 avian influenza. Infect Genet Evol. 2009;9:169–78. DOIPubMedGoogle Scholar
- Moscona A. Oseltamivir resistance—disabling our influenza defenses. N Engl J Med. 2005;353:2633–6. DOIPubMedGoogle Scholar
- World Health Organization Global Influenza Surveillance Network. Manual for the laboratory diagnosis and virological surveillance of influenza. 2011 [cited 2013 Aug 6]. http://whqlibdoc.who.int/publications/2011/9789241548090_eng.pdf
- World Health Organization/ World Organisation for Animal Health/ Food and Agriculture Organization (WHO/OIE/FAO) H5N1 Evolution Working Group. Continued evolution of highly pathogenic avian influenza A (H5N1): updated nomenclature. Influenza Other Respi Viruses. 2012;6:1–5 .
- Nguyen HT, Fry AM, Gubareva LV. Neuraminidase inhibitor resistance in influenza viruses and laboratory testing methods. Antivir Ther. 2012;17:159–73. DOIPubMedGoogle Scholar
- Deyde VM, Nguyen T, Bright RA, Balish A, Shu B, Lindstrom S, Detection of molecular markers of antiviral resistance in influenza A (H5N1) viruses using a pyrosequencing method. Antimicrob Agents Chemother. 2009;53:1039–47. DOIPubMedGoogle Scholar
- Nguyen HT, Sheu TG, Mishin VP, Klimov AI, Gubareva LV. Assessment of pandemic and seasonal influenza A (H1N1) virus susceptibility to neuraminidase inhibitors in three enzyme activity inhibition assays. Antimicrob Agents Chemother. 2010;54:3671–7. DOIPubMedGoogle Scholar
- Okomo-Adhiambo M, Sleeman K, Ballenger K, Nguyen HT, Mishin VP, Sheu TG, Neuraminidase inhibitor susceptibility testing in human influenza viruses: a laboratory surveillance perspective. Viruses. 2010;2:2269–89. DOIPubMedGoogle Scholar
- Gubareva LV, Trujillo AA, Okomo-Adhiambo M, Mishin VP, Deyde VM, Sleeman K, Comprehensive assessment of 2009 pandemic influenza A (H1N1) virus drug susceptibility in vitro. Antivir Ther. 2010;15:1151–9. DOIPubMedGoogle Scholar
- Triana-Baltzer GB, Gubareva LV, Klimov AI, Wurtman DF, Moss RB, Hedlund M, Inhibition of neuraminidase inhibitor-resistant influenza virus by DAS181, a novel sialidase fusion protein. PLoS ONE. 2009;4:e7838. DOIPubMedGoogle Scholar
- Klimov A, Balish A, Veguilla V, Sun H, Schiffer J, Lu X, Influenza virus titration, antigenic characterization, and serological methods for antibody detection. Methods Mol Biol. 2012;865:25–51. DOIPubMedGoogle Scholar
- Naughtin M, Dyason JC, Mardy S, Sorn S, von Itzstein M, Buchy P. Neuraminidase inhibitor sensitivity and receptor-binding specificity of Cambodian clade 1 highly pathogenic H5N1 influenza virus. Antimicrob Agents Chemother. 2011;55:2004–10. DOIPubMedGoogle Scholar
- World Health Organization. Meetings of the WHO working group on surveillance of influenza antiviral susceptibility– Geneva, November 2011 and June 2012. Wkly Epidemiol Rec. 2012;87:369–74 .PubMedGoogle Scholar
- Yen HL, Ilyushina NA, Salomon R, Hoffmann E, Webster RG, Govorkova EA. Neuraminidase inhibitor-resistant recombinant A/Vietnam/1203/04 (H5N1) influenza viruses retain their replication efficiency and pathogenicity in vitro and in vivo. J Virol. 2007;81:12418–26. DOIPubMedGoogle Scholar
- de Jong MD, Tran TT, Truong HK, Vo MH, Smith GJ, Nguyen VC, Oseltamivir resistance during treatment of influenza A (H5N1) infection. N Engl J Med. 2005;353:2667–72 . DOIPubMedGoogle Scholar
- Le QM, Kiso M, Someya K, Sakai YT, Nguyen TH, Nguyen KH, Avian flu: isolation of drug-resistant H5N1 virus. Nature. 2005;437:1108. DOIPubMedGoogle Scholar
- McKimm-Breschkin JL, Selleck PW, Usman TB, Johnson MA. Reduced sensitivity of influenza A (H5N1) to oseltamivir. Emerg Infect Dis. 2007;13:1354–7 .PubMedGoogle Scholar
- Stoner TD, Krauss S, DuBois M, Negovetich J, Stallknecht DE, Senne DA, Antiviral susceptibility of avian and swine influenza virus of the N1 neuraminidase subtype. J Virol. 2010;84:9800–9. DOIPubMedGoogle Scholar
- Kiso M, Ozawa M, Le MT, Imai H, Takahashi K, Kakugawa S, Effect of an asparagine-to-serine mutation at position 294 in neuraminidase on the pathogenicity of highly pathogenic H5N1 influenza A virus. J Virol. 2011;85:4667–72. DOIPubMedGoogle Scholar
- Collins PJ, Haire LF, Lin YP, Liu J, Russell RJ, Walker PA, Crystal structures of oseltamivir-resistant influenza virus neuraminidase mutants. Nature. 2008;453:1258–61. DOIPubMedGoogle Scholar
- Ilyushina NA, Seiler JP, Rehg JE, Webster RG, Govorkova EA. Effect of neuraminidase inhibitor-resistant mutations on pathogenicity of clade 2.2 A/Turkey/15/06 (H5N1) influenza virus in ferrets. PLoS Pathog. 2010;6:e1000933. DOIPubMedGoogle Scholar
- Earhart KC, Elsayed NM, Saad MD, Gubareva LV, Nayel A, Deyde VM, Oseltamivir resistance mutation N294S in human influenza A(H5N1) virus in Egypt. J Infect Public Health. 2009;2:74–80. DOIPubMedGoogle Scholar
- Amaro RE, Swift RV, Votapka L, Li WW, Walker RC, Bush RM. Mechanism of 150-cavity formation in influenza neuraminidase. Nat Commun. 2011;2:388.
- Hurt AC, Holien JK, Barr IG. In vitro generation of neuraminidase inhibitor resistance in A(H5N1) influenza viruses. Antimicrob Agents Chemother. 2009;53:4433–40. DOIPubMedGoogle Scholar
- Nguyen HT, Fry AM, Loveless PA, Klimov AI, Gubareva LV. Recovery of a multidrug-resistant strain of pandemic influenza A 2009 (H1N1) virus carrying a dual H275Y/I223R mutation from a child after prolonged treatment with oseltamivir. Clin Infect Dis. 2010;51:983–4. DOIPubMedGoogle Scholar
- Le MT, Wertheim HF, Nguyen HD, Taylor W, Hoang PV, Vuong CD, Influenza A H5N1 clade 2.3.4 virus with a different antiviral susceptibility profile replaced clade 1 virus in humans in northern Vietnam. PLoS ONE. 2008;3:e3339. DOIPubMedGoogle Scholar
- Chen H, Smith GJ, Li KS, Wang J, Fan XH, Rayner JM, Establishment of multiple sublineages of H5N1 influenza virus in Asia: implications for pandemic control. Proc Natl Acad Sci U S A. 2006;103:2845–50. DOIPubMedGoogle Scholar
- Boltz DA, Douangngeun B, Phommachanh P, Sinthasak S, Mondry R, Obert C, Emergence of H5N1 avian influenza viruses with reduced sensitivity to neuraminidase inhibitors and novel reassortants in Lao People’s Democratic Republic. J Gen Virol. 2010;91:949–59. DOIPubMedGoogle Scholar
- Wathen MW, Barro M, Bright RA. Antivirals in seasonal and pandemic influenza-future perspectives. Influenza Other Respitory Viruses. 2013;7:76–80.
- Chan-Tack KM, Gao A, Himaya AC, Thompson EG, Singer ME, Uyeki TM, Clinical experience with intravenous zanamivir under an emergency investigational new drug program in the United States. J Infect Dis. 2013;207:196–8. DOIPubMedGoogle Scholar
- White NJ, Webster RG, Govorkova EA, Uyeki TM. What is the optimal therapy for patients with H5N1 influenza? PLoS Med. 2009;6:e1000091. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 19, Number 12—December 2013
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Larisa V. Gubareva, Influenza Division, Centers for Disease Control and Prevention, Mailstop G16, 1600 Clifton Rd, Atlanta, GA, 30333, USA; .Larisa V. Gubareva, Centers for Disease Control and Prevention, 1600 Clifton Rd, Mailstop G16, Atlanta, GA, 30333, USA
Top