Volume 9, Number 5—May 2003
Research
Entamoeba moshkovskii Infections in Children in Bangladesh
Cite This Article
Citation for Media
Abstract
Entamoeba moshkovskii cysts are morphologically indistinguishable from those of the disease-causing species E. histolytica and the nonpathogenic E. dispar. Although sporadic cases of human infection with E. moshkovskii have been reported, the organism is considered primarily a free-living ameba. No simple molecular detection tool is available for diagnosing E. moshkovskii infections. We used polymerase chain reaction (PCR) to detect E. moshkovskii directly in stool. We tested 109 stool specimens from preschool children in Bangladesh by PCR; 17 were positive for E. histolytica (15.6%) and 39 were positive for E. dispar (35.8%). In addition, we found that 23 (21.1%) were positive for E. moshkovskii infection, and 17 (73.9%) of these also carried E. histolytica or E. dispar. The high association of E. moshkovskii with E. histolytica and E. dispar may have obscured its identification in previous studies. The high prevalence found in this study suggests that humans may be a true host for this ameba.
Entamoeba moshkovskii, considered to be primarily a free-living ameba, is indistinguishable in its cyst and trophozoite forms from E. histolytica (the cause of invasive amebiasis) and E. dispar (a common noninvasive parasite), except in cases of invasive disease when E. histolytica trophozoites may contain ingested red blood cells. E. moshkovskii has so far rarely been shown to infect humans; however, the organism appears to be ubiquitous in anoxic sediments. Although the early isolations of this species were from sewage, E. moshkovskii can also be found in environments ranging from clean riverine sediments to brackish coastal pools (1). E. moshkovskii is osmotolerant, can be cultured at room temperature, and is resistant to emetine, all characteristics that distinguish it from E. histolytica and E. dispar (2–5). Human isolates of E. moshkovskii to date have come from North America, Italy, South Africa, and Bangladesh, and they have never been associated with disease (5,6). However, few studies have actually set out to identify such infections (7).
The structural resemblance of the apparently innocuous E. moshkovskii to the disease-causing E. histolytica makes differentiating the two species important. In the clinical setting, for example, an E. moshkovskii–infected patient could be diagnosed as infected with E. histolytica and be treated unnecessarily with antiamebic chemotherapy. Most studies that have investigated the prevalence of E. histolytica and E. dispar have not considered the possible presence of E. moshkovskii, partly because of a lack of tools to detect E. moshkovskii other than cultivation, which is labor-intensive, not always successful, and problematic in the case of mixed infections. We report for the first time the application of tools to detect the species directly in stool and investigate the prevalence of E. moshkovskii in humans, a group of children in an E. histolytica–and E. dispar–endemic area where the first human infection with E. moshkovskii from Bangladesh was detected (6).
Stool Specimens
Fecal specimens included in this study were from 109 preschool children ages 2–5 years from Mirpur, an urban slum in Dhaka, Bangladesh. Based on results of polymerase chain reaction (PCR) on stool DNA samples, 39 were E. dispar–positive, 17 were E. histolytica–positive, and 1 was positive for both E. histolytica and E. dispar. Of the 52 samples negative by stool PCR, 18 were eventually found positive for E. histolytica, E. dispar, or both, either by PCR from culture DNA or by antigen detection tests performed on stool specimens, and the remaining 34 samples were negative by all methods. Only four of the samples were from children with diarrhea.
Cell Culture and Isoenzyme Analysis
All stool samples were cultured for Entamoeba species in Robinson’s medium (8) within 6 hours of collection, and hexokinase isoenzyme analysis was performed when possible as previously described (9). E. moshkovskii strains Laredo and FIC were maintained axenically in LYI-S-2 medium (10) with 10% adult bovine serum. Laredo (ATCC 30042) is a human isolate, and FIC (ATCC 30041) is an environmental isolate. E. histolytica HM-1:IMSS clone 9 (ATCC 50528) and E. dispar SAW760 (ATCC 50484) were used as controls.
Antigen Detection Tests for E. histolytica and E. dispar
The TECHLAB, Inc. (Blacksburg, VA) Entamoeba test (designed to detect but not differentiate E. histolytica and E. dispar antigen in stool specimens) and E. histolytica test (designed to detect specifically E. histolytica in stool specimens) were performed on stool specimens according to the manufacturer’s instructions (9).
Preparation of DNA
Stool DNA was isolated by using a modified version of the silica-DNA binding method of Katzwinkel-Wladarsch et al. as previously described (11,12). Culture DNA was isolated by a cetyltrimethylammonium bromide (CTAB) extraction method as previously described (13), dissolved in 10 mM Tris-Cl (pH 8.5), and passed over a Microspin S-200 HR column (Amersham Biosciences UK Ltd, Chalfont St. Giles, England). RNA was removed by the addition of RNase A (Promega UK, Ltd, Southampton, England) to 0.05 µg mL-1.
Small Subunit rRNA Gene Amplification
Based on the sequences of the small subunit rRNA genes (SSU-rDNA) of E. histolytica and E. dispar, nested sets of primers (designated E-1/E-2, Eh-1/Eh-2, and Ed-1/Ed-2) were used, as described (11), to detect E. histolytica and E. dispar in stool specimens (Table 1). Based on the sequence of the SSU-rDNA gene of E. moshkovskii Laredo (GenBank accession no. AF 149906), a nested set of primers (designated Em-1/Em-2 and nEm-1/nEm-2) was designed (unpub. data) and used to detect E. moshkovskii in stool DNA (Table 1). In the initial PCR (total vol. 25 µL), 1.0 µL of stool or culture DNA was used. Thermal cycler conditions included 30 cycles, each consisting of 92°C for 1 min, 55°C for 1 min, and 72°C for 1 min, followed by a final extension of 7 min at 72°C. In the nested PCR, 1.0 µL of first PCR product was used as the template DNA and the annealing temperature was raised to 62°C, leaving the other parameters of the amplification cycles unchanged. E. moshkovskii–specific nested SSU-rDNA gene amplification products were digested with restriction endonuclease XhoI for 1 h at 37°C according to the manufacturer’s instructions (Invitrogen Corp., Carlsbad, CA) to verify species identity. All PCR products were separated in 1.8% NuSieve 3:1 agarose gels (Flowgen, Lichfield, England) in 1x Tris-borate-EDTA buffer and visualized after staining with ethidium bromide (0.2 µg mL-1; Sigma-Aldrich Co. Ltd, Poole, England).
ArgTCT Gene PCR Amplification
Based on the sequence of the ArgTCT tRNA gene of E. histolytica, a set of primers were designed (ArgTCT-1 and ArgTCT-2). Thermal cycler conditions for PCR were the following: 30 cycles each, consisting of 94°C for 1 min, 55°C for 1 min 30 s, and 72°C for 2 min, followed by a final extension of 5 min at 72°C. The ArgTCT amplification products from E. moshkovskii Laredo, E. moshkovskii MS15-3646 (one of the infections detected above), and E. dispar SAW760 were cloned into the pGEM-T Easy vector (Promega) and sequenced (MWG Biotech Ltd, Milton Keynes, England). From the sequence results, an E. moshkovskii–specific primer pair, EmR-1 and EmR-2, was designed to amplify the E. moshkovskii ArgTCT gene fragment specifically (Table 1). PCR amplification was performed at an annealing temperature of 58°C as described for ArgTCT gene amplification.
Culture and Isoenzyme Analysis
All 109 stool specimens were added to Robinson’s medium for growth of Entamoeba species. Incubation led to growth of E. histolytica/E. dispar/E. moshkovskii in 33 cultures and E. coli in 8 cultures (no growth of E. hartmanni or Endolimax nana or was observed). Hexokinase isoenzyme analysis was possible for 10 cultures; 4 of them showed the band pattern of Entamoeba histolytica, 5 showed E. dispar, and 1 showed the band pattern of E. dispar with an extra band just behind the faster moving band, perhaps indicating a mixed culture with E. moshkovskii.
Detection of E. moshkovskii by Nested PCR
Figure 1
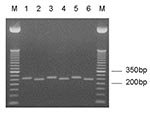
Figure 1. Entamoeba moshkovskii–specific nested SSU rDNA polymerase chain reaction (PCR) products. Odd- and even-numbered lanes represent undigested and XhoI-digested PCR products, respectively. Lanes 1/2, E. moshkovskii Laredo; lanes 3/4–5/6, DNA from stool...
The reference strain E. moshkovskii Laredo gave the expected band at approximately 260 bp with the E. moshkovskii–specific SSU-rDNA nested primers, whereas control E. histolytica HM-1:IMSS and E. dispar SAW760 DNAs were negative. Twenty-three of 109 (21%) stool DNA samples were positive by nested PCR for E. moshkovskii (Table 2). Of these, seven were positive for amebae by culture; one DNA sample extracted from these cultures was positive for E. moshkovskii. Seventeen of the 23 E. moshkovskii–positive samples were also positive for E. histolytica, E. dispar, or both, by either PCR of stool SSU rDNA (13/17) or by TECHLAB Entamoeba or E. histolytica tests (15/17) (Figure 1). One of the four children with diarrhea was positive for E. moshkovskii and coinfected with E. dispar. The cause of his diarrhea remained undetermined.
Figure 2

Figure 2. ArgTCT locus. ArgTCT sequences from Entamoeba histolytica HM-1:IMSS (GenBank accession no. AZ535059), E. dispar SAW760 (GenBank accession no. AF 525284), E. moshkovskii Laredo (GenBank accession no. AF 525285), and MS15-3646 (GenBank...
A comparison of SSU-rDNA sequences from E. moshkovskii, E. histolytica, and E. dispar, showed that the restriction endonuclease XhoI cuts exclusively in the E. moshkovskii–specific, 258-bp–nested PCR product to produce 236-bp and 22-bp fragments. Products from all 23 positive stool samples and the Laredo strain showed the presence of this site (Figure 2).
ArgTCT PCR and Sequence Analysis
To detect polymorphism among the E. moshkovskii samples, we studied a locus known to show polymorphism in E. histolytica and E. dispar (unpub. data). The ArgTCT primers amplify E. histolytica, E. dispar, and E. moshkovskii DNA. The sizes of the PCR products from E. histolytica HM-1:IMSS, E. dispar SAW760, and E. moshkovskii Laredo were 586 bp, 586 bp, and 323 bp, respectively. We did not observe a band in the 250- to 350-bp region in any of the E. histolytica and E. dispar strains with these primers (data not shown). Because 17 of 23 E. moshkovskii–positive samples were also positive for E. histolytica, E. dispar (by SSU-rDNA PCR or TECHLAB enzyme-linked immunosorbent assay), or both, we ignored products in the 500- to 600-bp region (assuming that they were derived from E. histolytica or E. dispar DNA) and considered a sample positive for E. moshkovskii when it produced a band at approximately 300 bp. By this criterion, we found 18 of 23 samples were positive for E. moshkovskii, and they showed slight PCR product size variation (data not shown). The PCR products from one stool sample, E. moshkovskii Laredo and E. dispar SAW760, were cloned, sequenced, and aligned with that of E. histolytica HM-1:IMSS, and E. moshkovskii–specific primers (EmR-1 and EmR-2) were designed (Figure 2) In addition to notable PCR product size differences, analysis clearly showed that the E. moshkovskii sequence is completely different from those of E. histolytica and E. dispar and, unlike the E. histolytica and E. dispar sequences, it contains no short tandem repeat sequences (Figure 2C).
Figure 3
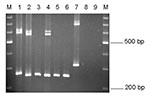
Figure 3. EmR polymerase chain reaction products. Lane 1, Entamoeba moshkovskii Laredo; lane 2, E. moshkovskii ICDDRB:717; lanes 3–6, E. moshkovskii–positive stool DNA samples; lane 7, E. moshkovskii FIC; lane 8, E. histolytica...
The EmR primers amplified the expected 265-bp fragment from E. moshkovskii Laredo DNA and did not amplify E. histolytica HM-1:IMSS or E. dispar SAW760 DNA. However, they successfully amplified 10 of a possible 23 E. moshkovskii–positive stool DNA samples. The most likely reason why these primers did not amplify the other 13 E. moshkovskii DNA samples is that they differed in sequence in the primer-binding regions. Although the PCR product size of the 10 positive samples was slightly different from that of Laredo, they were very similar in size to each other (Figure 3). The DNA of the previously reported E. moshkovskii ICDDRB:717, isolated from humans in the same geographic location (6), also gave a product of the same size (Figure 3, lane 2). The EmR primers successfully amplified DNA from environmental E. moshkovskii isolate FIC, but its product size was quite different from that of the human isolates of E. moshkovskii (Figure 3, lane 7).
The main objectives of this study were to develop molecular tools to identify E. moshkovskii and to investigate its prevalence and diversity in humans. We were successful in developing a simple diagnostic technique: a nested SSU-rDNA PCR followed by restriction endonuclease digestion. We chose to use nested PCR to detect E. moshkovskii infections because our previous experience in this area showed that nested PCR was much more efficient in amplifying stool DNA (14). Our attempt to produce a species-specific polymorphic marker was not completely successful. The EmR primers failed to amplify 13 of 23 E. moshkovskii–containing samples, probably because of sequence differences in primer-binding sites. However, the ArgTCT primers, originally designed to amplify E. histolytica and E. dispar DNA, did amplify most of the E. moshkovskii samples, producing a product distinct in size from those of E. histolytica and E. dispar.
Our study has some limitations. The subjects were children 2–5 years of age, so we do not know whether these subjects are representive of all age groups. All previous human isolates of E. moshkovskii have belonged to ribodeme 2 (5). Our attempts to perform riboprinting on these infections were unsuccessful, likely because of the size of the amplification target (approximately 1.95 kb). Even if PCR had been successful, the presence of mixed infections with other eukaryotes would have prevented successful typing.
This study has several important findings. The overall E. moshkovskii prevalence (21%) suggests that this infection is common among these children. E. dispar–infected children were almost twice as likely to have a mixed infection with E. moshkovskii (35%) compared to those with (18%) or without E. histolytica (18%) infections. None of the six children with E. moshkovskii monoinfections had diarrhea or dysentery, which suggests that E. moshkovskii is a noninvasive parasite. The high prevalence of E. moshkovskii infection may have been unnoticed over the years because most such infections (74%) were mixed infections with E. histolytica, E. dispar, or both. Previous attempts to identify human E. moshkovskii infections (7) may have failed because the human intestinal flora was unsuitable for cultivation at room temperature.
The high prevalence of E. moshkovskii shown in this study population indicates that perhaps humans are a true host for this putatively free-living ameba and are not just transiently infected. This prevalence may also explain some of the microscopy- positive/antigen-negative results obtained when using the Entamoeba test kit (15). Epidemiologic studies of E. histolytica infection should include tools to diagnose all three of these species individually, simultaneously, and accurately, and the prevalence of E. moshkovskii infection in other regions of the world should be investigated.
Mr. Ali is a senior research officer at the International Centre for Diarrheal Disease Research, Dhaka, Bangladesh, on leave and studying for his doctorate in the Department of Infectious and Tropical Diseases, London School of Hygiene and Tropical Medicine. His current research focus includes genetic diversity in Entamoeba species.
Mr. Hossain is a research officer at the International Centre for Diarrheal Disease Research, Dhaka, Bangladesh. His interests include the diagnosis of Entamoeba species and parasite-related nutritional biochemistry.
Acknowledgment
This research was supported in part by a grant received by International Centre for Diarrheal Disease Research, Dhaka, Bangladesh, Centre for Health and Population Research, with the support of University of Virginia (NIH grant AI-43596) and by grant 067314 from the Wellcome Trust awarded to C.G. Clark. The International Centre for Diarrheal Disease Research, Dhaka, Bangladesh, acknowledges with gratitude the commitment of University of Virginia to the Centre’s research efforts. Dr. Ali is funded by the Commonwealth Scholarship Commission.
References
- Clark CG, Diamond LS. Intraspecific variation and phylogenetic relationships in the genus Entamoeba as revealed by riboprinting. J Eukaryot Microbiol. 1997;44:142–54. DOIPubMedGoogle Scholar
- Dreyer DA. Growth of a strain of Entamoeba histolytica at room temperature. Tex Rep Biol Med. 1961;19:393–6.PubMedGoogle Scholar
- Entner N, Most H. Genetics of Entamoeba: characterization of two new parasitic strains that grow at room temperature (and at 37°C). J Protozool. 1965;12:10–3.PubMedGoogle Scholar
- Richards CS, Goldman M, Cannon LT. Cultivation of Entamoeba histolytica and Entamoeba histolytica-like strains at reduced temperature and behavior of the amebae in diluted media. Am J Trop Med Hyg. 1966;5:648–55.PubMedGoogle Scholar
- Clark CG, Diamond LS. The Laredo strain and other Entamoeba histolytica–like amoebae are Entamoeba moshkovskii. Mol Biochem Parasitol. 1991;46:11–8. DOIPubMedGoogle Scholar
- Haque R, Ali IKM, Clark CG, Petri WA Jr. A case report of Entamoeba moshkovskii infection in a Bangladeshi child. Parasitol Int. 1998;47:201–2. DOIGoogle Scholar
- Robinson GL, Sargeaunt PG. Low temperature strains of Entamoeba histolytica. Trans R Soc Trop Med Hyg. 1969;63:412–3. DOIPubMedGoogle Scholar
- Robinson GL. The laboratory diagnosis of human parasitic amoebae. Trans R Soc Trop Med Hyg. 1968;62:285–94. DOIPubMedGoogle Scholar
- Haque R, Kress K, Wood S, Jackson TFHG, Lyerly D, Wilkins T, Diagnosis of pathogenic Entamoeba histolytica infection using a stool ELISA based on monoclonal antibodies to the galactose-specific adhesin. J Infect Dis. 1993;167:247–9.PubMedGoogle Scholar
- Clark CG, Diamond LS. Methods for cultivation of luminal parasitic protists of clinical importance. Clin Microbiol Rev. 2002;15:329–41. DOIPubMedGoogle Scholar
- Haque R, Ali IKM, Akther S, Petri WA Jr. Comparison of PCR, isoenzyme analysis, and antigen detection for diagnosis of Entamoeba histolytica infection. J Clin Microbiol. 1998;36:449–52.PubMedGoogle Scholar
- Katzwinkel-Wladarsch S, Löscher T, Rinder H. Direct amplification and differentiation of pathogenic and nonpathogenic Entamoeba histolytica DNA from stool specimens. Am J Trop Med Hyg. 1994;51:115–8.PubMedGoogle Scholar
- Clark CG, Diamond LS. Entamoeba histolytica: a method for isolate identification. Exp Parasitol. 1993;7:450–5. DOIPubMedGoogle Scholar
- Ayeh-Kumi PF, Ali IKM, Lockhart LA, Gilchrist CA, Petri WA Jr, Haque R. Entamoeba histolytica: genetic diversity of clinical isolates from Bangladesh as demonstrated by polymorphisms in the serine-rich gene. Exp Parasitol. 2001;99:80–8. DOIPubMedGoogle Scholar
- Haque R, Neville LM, Hahn P, Petri WA Jr. Rapid diagnosis of Entamoeba infection by using Entamoeba and Entamoeba histolytica stool antigen detection kits. J Clin Microbiol. 1995;33:2558–61.PubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 9, Number 5—May 2003
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
C. Graham Clark, Department of Infectious and Tropical Diseases, London School of Hygiene and Tropical Medicine, Keppel Street, London, WC1E 7HT, England; fax: +44-207-636-8739
Top