Volume 16, Number 10—October 2010
Research
Influenza A (H5N1) Viruses from Pigs, Indonesia
Cite This Article
Citation for Media
Abstract
Pigs have long been considered potential intermediate hosts in which avian influenza viruses can adapt to humans. To determine whether this potential exists for pigs in Indonesia, we conducted surveillance during 2005–2009. We found that 52 pigs in 4 provinces were infected during 2005–2007 but not 2008–2009. Phylogenetic analysis showed that the viruses had been introduced into the pig population in Indonesia on at least 3 occasions. One isolate had acquired the ability to recognize a human-type receptor. No infected pig had influenza-like symptoms, indicating that influenza A (H5N1) viruses can replicate undetected for prolonged periods, facilitating avian virus adaptation to mammalian hosts. Our data suggest that pigs are at risk for infection during outbreaks of influenza virus A (H5N1) and can serve as intermediate hosts in which this avian virus can adapt to mammals.
A highly pathogenic avian influenza virus A (H5N1) was first recognized among geese in Guangdong Province, southern People’s Republic of China, in 1996 (1). Within a year, this goose virus underwent reassortment with viruses circulating in other avian species. By 1997, the virus had become widespread among poultry in Hong Kong, and direct avian-to-human transmission of influenza A (H5N1) viruses was reported (2,3). Since late 2003, influenza A (H5N1) viruses have spread to domestic poultry in other Southeast Asian countries (4). Since mid 2005, they have been detected across Asia, Europe, and Africa, causing severe damage to the poultry industry and infecting >490 humans, resulting in a mortality rate of 60% (5–8). Indonesia has been particularly affected by these viruses; >160 cases of human infection (i.e., about one third of the total confirmed human influenza A (H5N1) infections worldwide) and a mortality rate >80% have been reported (8). Hence, understanding prevalence and adaptation of influenza A (H5N1) influenza viruses in Indonesia is crucial.
Influenza viruses attach to host cells by binding their hemagglutinin (HA) to cell-surface oligosaccharides containing a terminal sialic acid. The HA of avian influenza viruses preferentially binds to sialic acid linked to galactose by α-2,3 linkages (SAα2,3Gal); that of human viruses binds to SAα2,6Gal (9). Correspondingly, epithelial cells in the upper respiratory tracts of humans mainly bear SAα2,6Gal receptors (10,11), and those in duck intestines (the major replication site for duck viruses) mainly possess SAα2,3Gal (12). Virus receptor specificities and expression patterns of receptors on host cells are thought to be major determinants of the host range restriction of influenza viruses (13). Thus, the recognition of human-type receptors by avian viruses appears to be necessary for these viruses to replicate in the upper respiratory tract and be transmitted efficiently from human to human. Given that influenza A (H5N1) viruses isolated from humans are not transmitted efficiently despite their ability to recognize human-type receptors (14), mutations in the polymerase and other viral genes may also be needed for replication of influenza A (H5N1) viruses in the upper respiratory tract (15).
Traditionally, pigs have been considered as “mixing vessels” (16–19) because they support replication of avian and human influenza viruses (17). Their tracheal epithelial cells reportedly bear SAα2,3Gal and SAα2,6Gal receptors (18). However, recent studies have shown that despite SAα2,3Gal and SAα2,6Gal receptors in pig respiratory tracts, SAα2,3Gal is found only in the smaller airways (bronchioli and alveoli) and not in the trachea (20,21). Kuchipudi et al. (22) found SAα2,3Gal and SAα2,6Gal receptors in the bronchi, bronchioli, and alveoli of chickens and ducks; however, SAα2,6Gal was dominant in chicken tracheal epithelium, and SAα2,3Gal, in duck trachea. Given that influenza A (H5N1) viruses have been transmitted directly from birds to humans, the central dogma of pigs as a mixing vessel may no longer stand. Moreover, under experimental conditions, pig susceptibility to infection with avian influenza A (H5N1) viruses is low (23). Nevertheless, the pandemic (H1N1) 2009 virus is a reassortant that originated from 4 genetically distinct viruses and appeared to be generated in pigs (24), suggesting their role in the generation of pandemic influenza viruses. Infection of pigs with influenza A (H5N1) viruses has been reported in Vietnam (25) and China (26); however, the infection status of pigs in Indonesia remains unknown. We, therefore, explored whether pigs in Indonesia had been infected with influenza A (H5N1) viruses and, if so, whether the viruses were transmitted multiple times and had acquired the ability to recognize human-type receptors.
Specimen Collection
Figure 1

Figure 1. Provinces in Indonesia (gray shading) where surveillance for influenza A (H5N1) virus in pigs was conducted during 2005–2009.
Virologic and serologic surveillance was conducted during 3 rainy seasons during 2005–2009: January–February 2005, October–February 2007, and, November 2008–April 2009. Nasal, fecal, and serum samples were collected from apparently healthy pigs in various districts of Indonesia (Figure 1). The nasal and fecal samples were injected into 10-day-old embryonated eggs, and the allantoic fluid was tested for hemagglutination. Hemagglutination-positive allantoic fluid was subjected to reverse transcription–PCR by using H5 HA–specific and N1 neuraminidase–specific primers; only positive samples were tested further. Serum was analyzed to estimate the prevalence of influenza virus A (H5N1) infection.
Cells and Virus Isolation
MDCK cells and an MDCK cell line that overexpresses the human β-galactoside α-2,6-sialyltransferase I gene (MDCK-ST6GalI) (27) were maintained in minimal essential medium (MEM) containing 5% newborn calf serum at 37°C in 5% CO2. Virus isolation from specimens was performed by using 10-day-old embryonated chicken eggs, MDCK cells, or MDCK-ST6GalI cells in MEM containing 0.3% bovine serum albumin (BSA) (Sigma-Aldrich, Inc., St. Louis, MO, USA) (Table A1). Viruses isolated in MDCK cells were used whenever they were available.
Serologic Analysis
Swine serum samples were tested for neutralizing antibodies against influenza A/swine/East Java/UT6040/2007 (H5N1) and A/duck/Czechoslovakia/56 (H4N6) viruses. Subtype H4N6 was used as a negative control. Serum was mixed with 3 volumes of receptor-destroying enzyme (Denka Seiken Co., Ltd, Tokyo, Japan) overnight at 37°C and inactivated at 56°C for 30 min. A 2-fold serial dilution series of serum (1:4–1:512) was mixed with an equal volume of influenza virus at 100 TCID50 (50% tissue culture infectious doses) and incubated at 37°C for 30 min. Viruses were inoculated to monolayers of MDCK cells for 1 h, washed 2×, and incubated with MEM containing 0.3% BSA for 2 d at 37°C in a 5% CO2 incubator. Cytopathic effects were observed to determine the neutralizing activity of the test serum. The detection limit for the neutralizing antibody was <4 dilutions of serum.
Sequence Analysis
To characterize the swine influenza A (H5N1) viruses isolated in Indonesia, we sequenced the HA genes of 39 viruses isolated from pigs in Banten, East Java, North Sumatra, and South Kalimantan provinces and grouped them according to their genetic similarities. Viral RNA was extracted with ISOGEN (Nippon Gene, Tokyo, Japan) according to the manufacturer’s instructions. Extracted RNA was reverse transcribed with SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA, USA) and an oligonucleotide complementary to the 12-nt sequence at the 3′ end of the viral RNA and amplified by PCR with Pfu-ultra (Stratagene, La Jolla, CA, USA) or Phusion (Finnzymes, Espoo, Finland) high-fidelity DNA polymerase and primers specific for each segment of the influenza virus A (H5N1). Primer sequences are available upon request. The PCR products were separated by agarose gel electrophoresis, purified by using a MinElute Gel Extraction Kit (QIAGEN, Hilden, Germany), and then sequenced. The nucleotide sequences obtained in this study are available from GenBank, accession nos. HM440051–HM440154.
Phylogenetic Analysis
We phylogenetically analyzed 13 representative swine influenza A (H5N1) viruses for all 8 viral genes and compared these sequences with publicly available sequences. All sequences were assembled and edited with BioEdit 7 software (28). Neighbor-joining tree analysis was conducted by using ClustalW (www.clustal.org). Estimates of the phylogenies were calculated by performing 100 neighbor-joining bootstrap replicates.
Receptor Specificity Assays
During replication in pigs, avian influenza viruses may adapt to recognize human-type receptors because such receptors are present in the epithelial cells of pig trachea (18). We therefore analyzed the receptor specificity of representative viruses from each of the 3 swine groups: A/swine/Banten/UT3081/2005 for the 2005 swine group, A/swine/East Java/UT6012/2007 for the 2006–07 swine (A) group, and A/swine/Banten/UT6001/2006 for the 2006–07 swine (B) group. We also analyzed A/swine/Banten/UT3062/2005 clone 6 and A/swine/Banten/3063/05 clone 1, each of which possesses a single amino acid change in HA that distinguishes it from other clones. The receptor specificity of these influenza A (H5N1) viruses was determined by use of an assay that measures direct binding to sialylglycopolymers possessing either SAα2,3Gal or SAα2,6Gal. We used this solid-phase binding assay with the sodium salts of sialylglycopolymers (poly α-
Virus Prevalence
Of 702 nasal swabs, 52 (7.4%) collected in 2005–2007 yielded influenza A (H5N1) viruses (Table 1; Table A1); no virus was isolated from fecal samples of the same pigs. All 35 viruses isolated in 2005 were from 5 pig farms in the Tangerang District of Banten Province, near an area in which an influenza A (H5N1) outbreak among poultry had been confirmed in 2004 (31) and where the virus has since remained enzootic. Samples collected from a slaughterhouse in the Surabaya district of East Java Province were negative for influenza virus A (H5N1). In the subsequent surveillance period, October 2006–February 2007, we detected viruses in pigs on 4 farms in the Tangerang, Kediri, and Medan districts of Banten, East Java, and North Sumatra, respectively, and in slaughterhouses in the Surabaya and Banjarmasin districts of East Java and South Kalimantan; all sites were near previous outbreaks of influenza virus A (H5N1) infection among poultry. Pigs from which these viruses were isolated did not show any signs of influenza-like illness at the time of sample collection. During the November 2008–April 2009 surveillance period, virus was not isolated from any nasal swabs from 300 pigs tested. However, the 300 serum samples tested indicated that 3 (1%) pigs had neutralizing antibodies against a subtype H5N1 virus but not subtype H4N6, suggesting limited exposure to influenza A (H5N1) viruses. These positive samples were obtained from a farm in the Malang District of East Java Province; neutralizing titers were 4–16 (Table 1).
Virus Sequences
Among the 39 viruses sequenced, the first group comprised 24 isolates collected in Banten Province during January 2005–February 2005; the HA genes of these viruses were either identical or differed by no more than 2 nt. The second group comprised 9 isolates collected during October 2006–February 2007 and also differed by only 2 nt, although the viruses were collected in different provinces: Banten, East Java, North Sumatra, and South Kalimantan. The HA genes in these 2 groups differed from each other by 49–53 nt. The third group included 6 isolates collected in Banten, East Java, and North Sumatra during October 2006–February 2007; these HAs were identical except for 1 nt and differed from those of the first and second groups by 42–45 and 58–61 nt, respectively. Thus, the swine influenza A (H5N1) viruses collected in our surveillance study could be classified into 3 distinct groups on the basis of their HA gene sequences, irrespective of the province from which they were isolated, suggesting extensive movement of pigs among provinces.
Phylogeny
Figure 2

Figure 2. Phylogenetic relationships among the A) hemagglutinin (HA) and B) neuraminidase (NA) genes of influenza A (H5N1) viruses isolated in Indonesia. The numbers below or above the branch nodes indicate neighbor-joining bootstrap...
Figure A1
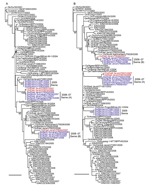
Figure A1. Phylogenetic relationships of the polymerase basic protein (PB) 2 (A) and PB1 (B) genes of influenza A (H5N1) viruses in Indonesia. All trees were generated by neighbor-joining in ClustalW (www.clustal.org)....
Figure A2
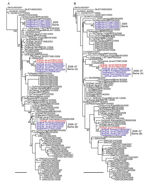
Figure A2. Phylogenetic relationships of the polymerase acidic protein (PA) (A) and nucleocapsid protein (NP) (B) genes of influenza A (H5N1) viruses in Indonesia. All trees were generated by neighbor-joining in ClustalW (<>
Figure A3
Figure A3. Phylogenetic relationships of the M (A) and NS (B) genes of H5N1 influenza viruses in Indonesia. All trees were generated by the neighbor-joining method in ClustalW (www.clustal.org). Numbers above or...
Phylogenetic analysis of the HA genes of the 13 representative viruses identified the same 3 groups described above. The HA genes of 4 viruses isolated in 2005 (2005 swine group) were placed in clade 2.1.1, and of the remaining 9 swine viruses isolated during 2006–2007, five were classified into the IDN/6/05-like sublineage (2006–07 swine [A] group) and 4 into clade 2.1.3 (2006–07 swine [B] group) (Figure 2, panel A). The most closely related strains of each swine virus group were chicken influenza A (H5N1) viruses: A/chicken/Indonesia/R60/2005 for the 2005 swine group, A/chicken/East Java/UT6016/2006 and A/chicken/East Java/UT6031/2007 for the 2006–07 swine (A) group, and A/chicken/East Java/UT6044/2007 for the 2006–07 swine (B) group. Analyses of the other 7 genes demonstrated that the phylogenetic relationships established for the HA gene were maintained; that is, the swine viruses in each group possessed nearly identical genes, and each group of swine viruses was most closely related to a chicken virus isolated near the site where the swine viruses were collected (Figure 2, panel B; Figure A1, Figure A2, Figure A3). Our results suggest that influenza A (H5N1) viruses were transmitted from avian species to pigs on at least 3 occasions.
Receptor Specificity
Figure 3
Figure 3. Receptor-binding activity of influenza A (H5N1) viruses. Direct binding of viruses to sialylglycopolymers containing either α2,3-linked (blue) or α2,6-linked (red) sialic acids was measured. A) Human isolate A/Kawasaki/173/2001; B) avian isolate...
Sequence analysis of the PCR products of the HA genes of A/swine/Banten/UT3062/2005 and A/swine/Banten/UT3063/2005 indicated that nucleotides were heterogeneous at certain positions, prompting us to plaque purify the viruses in MDCK cells to obtain viral clones with distinct HA sequences (Table 2). We found that most of the swine influenza subtype H5N1 isolates bound to only SAα2,3Gal, whereas the plaque-purified clone 6 of A/swine/Banten/UT3062/05 bound to SAα2,3Gal and SAα2,6Gal (Figure 3), indicating that during their replication in pigs, avian influenza A (H5N1) viruses can acquire the ability to recognize human virus receptors.
In contrast to the few reported cases of infection of pigs with highly pathogenic avian influenza A (H5N1) viruses (17,25,26), our surveillance study of 7 provinces in Indonesia during 3 periods shows that 7.4% of pigs surveyed during 2005–2007, but not 2008–2009, were infected with influenza A (H5N1) viruses. Phylogenetic analysis indicated that the viruses were transmitted to pigs on several different occasions, probably from poultry on nearby farms. According to the most recent classification of the HA gene (32,33), all avian and human influenza A (H5N1) viruses isolated in Indonesia belong to clade 2.1, which includes 3 well-defined lineages (clades 2.1.1–2.1.3) and a developing lineage termed IDN/6/05-like sublineage. In our study, all 24 viruses isolated during the first surveillance period belonged to the same cluster in clade 2.1.1 (2005 swine group) on the basis of recent HA classification (32,33). The 9 viruses collected during the second surveillance period belonged exclusively to the IDN/6/05-like sublineage, and the 6 remaining viruses collected during the same season were classified into clade 2.1.3; 2006–07 swine (A) and 2006–07 swine (B) groups, represented, respectively, by A/swine/East Java/UT6012/2007 and A/swine/Banten/UT6001/2006. Although no virus was isolated during the third surveillance period, 2008–09, a total of 3 (1%) pigs had neutralizing antibodies against influenza virus A (H5N1). These findings show that although influenza A (H5N1) viruses may not have been extensively circulating in pigs in Indonesia recently, these animals are susceptible to influenza A (H5N1) viruses and can serve as asymptomatic reservoirs for these viruses.
Because the phylogenetic relationships established for the HA gene extended to all viral genes, we conclude that the 3 groups of viruses identified in this survey were likely established independently, suggesting at least 3 separate avian-to-pig episodes of transmission of influenza A (H5N1) viruses during 2005–2009 in Indonesia. Our findings confirm sporadic reports of the susceptibility of pigs to influenza A virus (H5N1) infection in natural (25,26) and experimental settings (23,34) and suggest that when an outbreak of influenza A virus (H5N1) infection occurs on poultry farms, pigs on nearby farms should be evaluated for infection.
We also found evidence of pig-to-pig transmission of influenza A virus (H5N1), particularly among animals sampled during the first surveillance period. Many viruses possessing almost identical genes were isolated from pigs on the same farms (Table 1, Table 2). Pig-to-pig transmission would likely prolong the duration of influenza A (H5N1) virus infection within a pig population, thereby increasing the likelihood of adaptation and the subsequent generation of influenza A (H5N1) viruses that replicate efficiently in humans.
The lack of influenza-like signs in pigs infected with influenza A (H5N1) viruses has several public health implications. In Indonesia, pigs are transported to different locations according to market needs. This movement is reflected in our finding that clusters of swine viruses collected after 2006 were not consistent with those common to the sampling region. Indeed, viruses collected in North Sumatra, South Kalimantan, East Java, and Banten provinces showed identical or nearly identical genes, indicative of extensive transport of infected pigs throughout Indonesia. Thus, pathogenic influenza A (H5N1) viruses could easily evade detection as they spread through Indonesia in asymptomatic pigs being transported from province to province.
Our analysis of viral receptor specificities showed that 1 plaque-purified clone of A/swine/Banten/UT3062/2005 bound to avian-type and human-type receptors. Serine at position 134 was responsible for the human-type receptor recognition. This position is located within the 130-loop structural component of the receptor-binding pocket (35). Hence, the amino acid change at this position may affect receptor binding. Because serine at position 134 is never seen in avian influenza A (H5N1) viruses (alanine is highly conserved at this position in avian influenza A [H5N1] viruses), the Ala134Ser mutation probably occurred during adaptation of the virus to pigs. According to a previous report (36), human isolates possessing valine at this position could also bind to the human-type receptor, although a mutation at position 129 (L129V) was also required for the human-type receptor recognition in this strain. Therefore, mutations at position 134 probably correlated with human-type receptor recognition and may serve as molecular markers for assessing the pandemic potential of influenza virus A (H5N1) isolates.
Although influenza virus A (H5N1) infection was not reported among swine workers in Indonesia while we were collecting our pig specimens, a previous cohort study showed that such workers, as well as their unexposed spouses, had increased levels of antibody to swine influenza A (H1N1) viruses (37), suggesting that humans are indeed susceptible to swine-adapted viruses (38). The recent swine-origin pandemic (H1N1) 2009 further demonstrates that pigs can be a potential source of virus capable of causing a human influenza pandemic (24). These findings suggest that as influenza A (H5N1) viruses spread among pigs and adapt to recognize human-type receptors, farmers, swine workers, and their families will be at greatest risk for infection by the newly adapted viruses.
In summary, we found that influenza A (H5N1) viruses have been transmitted multiple times to pig populations in Indonesia and that 1 virus has acquired the ability to recognize human-type receptors. Of particular concern is that pigs infected with influenza A (H5N1) viruses showed no significant influenza-like signs and were likely transported to and from different provinces in Indonesia. On the basis of our findings, we encourage the Indonesian government to control the transport of pigs within Indonesia. Otherwise, opportunities for this avian virus to adapt to mammals will increase, as will the risk for emergence of a new pandemic influenza virus.
Dr Nidom is a director of the Faculty of Veterinary Medicine and the Collaborating Research Center–Emerging and Reemerging Infectious Diseases, Tropical Disease Centre, Airlangga University, Surabaya, Indonesia. His research interests include the pathobiology and epidemiology of poultry pathogens and zoonoses transmitted by poultry.
Acknowledgments
We thank Susan Watson for editing the manuscript.
This work was supported by a grant-in-aid for Specially Promoted Research, Japan; a contract research fund for the Program of Funding Research Centers for Emerging and Reemerging Infectious Diseases from the Ministries of Education, Culture, Sports, Science, and Technology, Japan; grants-in-aid of Health, Labor, and Welfare, Japan; an Exploratory Research for Advanced Technology grant from Japan Science and Technology Agency, Japan; and by National Institute of Allergy and Infectious Diseases Public Health Service research grants, USA.
References
- Xu X, Subbarao K, Cox NJ, Guo Y. Genetic characterization of the pathogenic influenza A/goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of influenza (H5N1) viruses from the 1997 outbreaks in Hong Kong. Virology. 1999;261:15–9. DOIPubMedGoogle Scholar
- Claas EC, Osterhaus AD, van Beek R, De Jong JC, Rimmelzwaan GF, Senne DA, Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet. 1998;351:472–7. DOIPubMedGoogle Scholar
- Subbarao K, Klimov A, Katze J, Regnery H, Lim W, Hall H, Characterization of an avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness. Science. 1998;279:393–6. DOIPubMedGoogle Scholar
- Li KS, Guan Y, Wang J, Smith GJ, Xu KM, Duan L, Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature. 2004;430:209–13. DOIPubMedGoogle Scholar
- Chen H, Smith GJ, Zhang SY, Qin K, Wang J, Li KS, Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature. 2005;436:191–2. DOIPubMedGoogle Scholar
- Ducatez MF, Olinger CM, Owoade AA, De Landtsheer S, Ammerlaan W, Niesters HG, Avian flu: multiple introductions of H5N1 in Nigeria. Nature. 2006;442:37. DOIPubMedGoogle Scholar
- Weber S, Harder T, Starick E, Beer M, Werner O, Hoffmann B, Molecular analysis of highly pathogenic avian influenza virus of subtype H5N1 isolated from wild birds and mammals in northern Germany. J Gen Virol. 2007;88:554–8. DOIPubMedGoogle Scholar
- World Health Organization. Cumulative number of confirmed human cases of avian influenza A/(H5N1) reported to WHO, 30 March 2010 [cited 2010 Apr 3]. http://www.who.int/csr/disease/avian_influenza/country/cases_table_2010_03_30/en/index.html
- Matrosovich MN, Gambaryan AS, Teneberg S, Piskarev VE, Yamnikova SS, Lvov DK, Avian influenza A viruses differ from human viruses by recognition of sialyloligosaccharides and gangliosides and by a higher conservation of the HA receptor–binding site. Virology. 1997;233:224–34. DOIPubMedGoogle Scholar
- Gambaryan A, Tuzikov A, Pazynina G, Bovin N, Balish A, Klimov A. Evolution of the receptor-binding phenotype of influenza A (H5) viruses. Virology. 2006;344:432–8. DOIPubMedGoogle Scholar
- Shinya K, Ebina M, Yamada S, Ono M, Kasai N, Kawaoka Y. Avian flu: influenza virus receptors in the human airway. Nature. 2006;440:435–6. DOIPubMedGoogle Scholar
- Matrosovich MN, Matrosovich TY, Gray T, Roberts NA, Klenk HD. Human and avian influenza viruses target different cell types in cultures of human airway epithelium. Proc Natl Acad Sci U S A. 2004;101:4620–4. DOIPubMedGoogle Scholar
- Suzuki Y, Ito T, Suzuki T, Holland RE Jr, Chambers TM, Kiso M, Sialic acid species as a determinant of the host range of influenza A viruses. J Virol. 2000;74:11825–31. DOIPubMedGoogle Scholar
- Yamada S, Suzuki Y, Suzuki T, Le MQ, Nidom CA, Sakai-Tagawa Y, Haemagglutinin mutations responsible for the binding of H5N1 influenza A viruses to human-type receptors. Nature. 2006;444:378–82. DOIPubMedGoogle Scholar
- Hatta M, Hatta Y, Kim JH, Watanabe S, Shinya K, Nguyen T, Growth of H5N1 influenza A viruses in the upper respiratory tracts of mice. PLoS Pathog. 2007;3:1374–9. DOIPubMedGoogle Scholar
- Castrucci MR, Donatelli I, Sidoli L, Barigazzi G, Kawaoka Y, Webster RG. Genetic reassortment between avian and human influenza A viruses in Italian pigs. Virology. 1993;193:503–6. DOIPubMedGoogle Scholar
- Kida H, Ito T, Yasuda J, Shimizu Y, Itakura C, Shortridge KF, Potential for transmission of avian influenza viruses to pigs. J Gen Virol. 1994;75:2183–8. DOIPubMedGoogle Scholar
- Ito T, Couceiro JN, Kelm S, Baum LG, Krauss S, Castrucci MR, Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J Virol. 1998;72:7367–73.PubMedGoogle Scholar
- Scholtissek C, Burger H, Kistner O, Shortridge KF. The nucleoprotein as a possible major factor in determining host specificity of influenza H3N2 viruses. Virology. 1985;147:287–94. DOIPubMedGoogle Scholar
- Nelli RK, Kuchipudi SV, White GA, Perez BB, Dunham SP, Chang KC. Comparative distribution of human and avian type sialic acid influenza receptors in the pig. BMC Vet Res. 2010;6:4. DOIPubMedGoogle Scholar
- Van Poucke SG, Nicholls JM, Nauwynck HJ, Van Reeth K. Replication of avian, human and swine influenza viruses in porcine respiratory explants and association with sialic acid distribution. Virol J. 2010;7:38. DOIPubMedGoogle Scholar
- Kuchipudi SV, Nelli R, White GA, Bain M, Chang KC, Dunham S. Differences in influenza virus receptors in chickens and ducks: implications for interspecies transmission. J Mol Genet Med. 2009;3:143–51.PubMedGoogle Scholar
- Lipatov AS, Kwon YK, Sarmento LV, Lager KM, Spackman E, Suarez DL, Domestic pigs have low susceptibility to H5N1 highly pathogenic avian influenza viruses. PLoS Pathog. 2008;4:e1000102. DOIPubMedGoogle Scholar
- Smith GJ, Vijaykrishna D, Bahl J, Lycett SJ, Worobey M, Pybus OG, Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature. 2009;459:1122–5. DOIPubMedGoogle Scholar
- Choi YK, Nguyen TD, Ozaki H, Webby RJ, Puthavathana P, Buranathal C, Studies of H5N1 influenza virus infection of pigs by using viruses isolated in Vietnam and Thailand in 2004. J Virol. 2005;79:10821–5. DOIPubMedGoogle Scholar
- Zhu Q, Yang H, Chen W, Cao W, Zhong G, Jiao P, A naturally occurring deletion in its NS gene contributes to the attenuation of an H5N1 swine influenza virus in chickens. J Virol. 2008;82:220–8. DOIPubMedGoogle Scholar
- Hatakeyama S, Sakai-Tagawa Y, Kiso M, Goto H, Kawakami C, Mitamura K, Enhanced expression of an alpha2,6-linked sialic acid on MDCK cells improves isolation of human influenza viruses and evaluation of their sensitivity to a neuraminidase inhibitor. J Clin Microbiol. 2005;43:4139–46. DOIPubMedGoogle Scholar
- Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucl Acids Symp Ser. 1999;41:95–8.
- Shinya K, Hatta M, Yamada S, Takada A, Watanabe S, Halfmann P, Characterization of a human H5N1 influenza A virus isolated in 2003. J Virol. 2005;79:9926–32. DOIPubMedGoogle Scholar
- Totani K, Kubota T, Kuroda T, Murata T, Hidari KI, Suzuki T, Chemoenzymatic synthesis and application of glycopolymers containing multivalent sialyloligosaccharides with a poly (
l -glutamic acid) backbone for inhibition of infection by influenza viruses. Glycobiology. 2003;13:315–26. DOIPubMedGoogle Scholar - World Organisation for Animal Health. Highly pathogenic avian influenza in Indonesia; 2 February 2004 [cited 2010 Aug 3]. ftp://ftp.oie.int/infos_san_archives/eng/2004/en_040206v17n06.pdf
- Takano R, Nidom CA, Kiso M, Muramoto Y, Yamada S, Sakai-Tagawa Y, Phylogenetic characterization of H5N1 avian influenza viruses isolated in Indonesia from 2003–2007. Virology. 2009;390:13–21. DOIPubMedGoogle Scholar
- WHO/OIE/FAO H5N1 Evolution Working Group. Toward a unified nomenclature system for highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis. 2008;14:e1.PubMedGoogle Scholar
- Shortridge KF, Zhou NN, Guan Y, Gao P, Ito T, Kawaoka Y, Characterization of avian H5N1 influenza viruses from poultry in Hong Kong. Virology. 1998;252:331–42. DOIPubMedGoogle Scholar
- Skehel JJ, Wiley DC. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu Rev Biochem. 2000;69:531–69. DOIPubMedGoogle Scholar
- Auewarakul P, Suptawiwat O, Kongchanagul A, Sangma C, Suzuki Y, Ungchusak K, An avian influenza H5N1 virus that binds to a human-type receptor. J Virol. 2007;81:9950–5. DOIPubMedGoogle Scholar
- Gray GC, McCarthy T, Capuano AW, Setterquist SF, Olsen CW, Alavanja MC. Swine workers and swine influenza virus infections. Emerg Infect Dis. 2007;13:1871–8.PubMedGoogle Scholar
- Van Reeth K. Avian and swine influenza viruses: our current understanding of the zoonotic risk. Vet Res. 2007;38:243–60. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 16, Number 10—October 2010
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Yoshihiro Kawaoka, Division of Virology, Department of Microbiology and Immunology, Institute of Medical Science, University of Tokyo, Shirokanedai, Minato-ku, Tokyo 108-8639, Japan
Top