Volume 18, Number 12—December 2012
Dispatch
Arctic-like Rabies Virus, Bangladesh
Cite This Article
Citation for Media
Abstract
Arctic/Arctic-like rabies virus group 2 spread into Bangladesh ≈32 years ago. Because rabies is endemic to and a major public health problem in this country, we characterized this virus group. Its glycoprotein has 3 potential N-glycosylation sites that affect viral pathogenesis. Diversity of rabies virus might have public health implications in Bangladesh.
Rabies virus causes severe encephalitis in a wide range of mammals, including humans. Conservative estimates suggest that 55,000 persons worldwide die of rabies each year (1). Although the case-fatality rate in humans is 100%, rabies is preventable by vaccination. Bangladesh has the world’s third highest death rate for human rabies, an estimated 2,100 deaths per year (2). Dogs are the main reservoir of the virus and are responsible for spillover infections in humans (2). Therefore, dogs should be the principal target for successful rabies elimination.
With political will and solid global epidemiologic information, rabies elimination is possible. Molecular typing of circulating rabies viruses is necessary to identify and develop effective control measures, and to understand the spread of certain rabies virus variants and their incursion into new regions (3). For rabies elimination, this knowledge is needed for establishing cooperative approaches between neighboring countries to which the disease is endemic.
Bangladesh is one of several countries in which no molecular study has been conducted to identify types of rabies virus circulating within its boundaries. A lack of knowledge of phylogenetic relationships of Bangladesh rabies virus with viruses in other countries continues to hinder coordinated rabies control efforts in the region. This study was conducted to characterize rabies virus circulating in Bangladesh and to determine its relationship with viruses in neighboring countries to clarify its epidemiologic relationships, origin, and transmission dynamics.
Seven brain samples were collected from animals with suspected rabies in 3 districts of Bangladesh (Dhaka, Narayanganj, and Narshingdi) in 2010 (Table 1). A portion of brainstem was removed from each sample and preserved in TRizol (Invitrogen, Carlsbad, CA, USA) at –20°C. Total RNA was extracted from brain homogenate, cDNA was synthesized by using random hexamer primers, reverse transcription PCR was conducted to amplify gene fragments, and nucleotide sequencing of genes was performed (4).
Full-length nucleoprotein (N) and glycoprotein (G) gene sequences from samples were determined. Nucleotide identities of N and G genes were 98%–100%. Amino acid identities of N and G genes were 100% and 98%–100%, respectively. Complete genomic sequencing (11,928 nt) of strain BDR5 was also conducted.
Evolutionary analysis was performed by using full-length N gene. We created a maximum clade credibility phylogenetic tree using the Bayesian Markov chain Monte Carlo method available in BEAST version 1.6.1 (5). Analysis was conducted by using a relaxed (uncorrelated lognormal) molecular clock and a generalized time reversible + Γ + proportion invariant model (6). All chains were run for 90 million generations and sampled every 3,000 steps and an effective sample size >1,383 was obtained for all estimated parameters. Posterior densities were calculated with 10% burn-in and checked for convergence by using Tracer version 1.5 in BEAST.
The mean rate of nucleotide substitution estimated for the N gene was 2.3 × 104 substitutions/site/year (95% highest posterior density [HPD] 1.4–3.1 × 104 substitutions/site/year). This rate is consistent with that of a previous study (7). The phylogenetic tree showed that rabies viruses in Bangladesh belong to Arctic/Arctic-like group 2 (AAL2) (3) also known as Arctic-like-1 (8), in close association with the strain from Bhutan.
Figure 1

Figure 1. . . Bayesian maximum credibility tree showing genealogy of rabies virus obtained by analyzing nucleotide sequences of full nucleoprotein (N) gene sequences (1,350 nt), Bangladesh. Nodes indicate the mean age at...
Approximately 397.0 years ago (95% HPD 273.5–589.5 years), AAL and cosmopolitan rabies virus segregated from their most recent common ancestor (Figure 1). Approximately 225.6 years ago (95% HPD 157.4–324.2 years), AAL3 segregated. Approximately 187.4 years ago (95% HPD 129.0–271.9 years), AAL1 and AAL2 segregated. The AAL2 clade had a common progenitor that circulated ≈133.1 years ago (95% HPD 91.3–193.4 years), which has evolved into several different lineages. One lineage evolved 91.5 years ago (95% HPD 63.1–132.2 years) and currently circulates in Bangladesh, India, and Bhutan. Separate linages circulate in others countries in this region, including Iran, Nepal, Pakistan, and Afghanistan. AAL2 spread into central Bangladesh 32.3 years ago (95% HPD 18.4–50.6 years) in ≈1978 (95% HPD range 1958–1991).
Compared with the AAL2 strain from India (AY956319), BDR5 had several amino acid substitutions (Table 2). Sizes of their 2 genomes, leader RNA, trailer RNA, and intergenic regions were similar. The N-glycosylation site was predicted by using the NetNGlyc 1.0 server (www.cbs.dtu.dk/server/netnglyc). With the exception of BDR6, the G gene of all strains had potential glycosylation sites at position 37, 146, and 319.
Genetic analysis and phylogenetic studies can contribute to understanding the epidemiology of rabies virus in disease-endemic countries. Molecular analysis of animal rabies viruses showed that AAL2 appeared in central Bangladesh only 32 years ago. A close association between N genes sequences from rabies viruses in Bangladesh and Bhutan indicates that they originated from a common ancestor. If one considers the ease of human movement between countries, AAL2 most likely entered Bangladesh from India rather than from Bhutan.
Figure 2
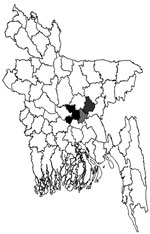
Figure 2. . . Three districts of Bangladesh from which samples were tested for Arctic/Arctic-like rabies virus and strains were found. Black, Dhaka District, strains BDR1, BDR3, and BDR6; light gray, Narayanganj District,...
Circumstantial evidence suggests that rabies virus spread from India to Bhutan (9). AAL2 circulates in many states of India. It has spread into southern India and has replaced older strains (10,11). It is likely that AAL2 is also circulating in states of India that are between Bhutan and Bangladesh. Estimated time of AAL2 spread is based on 7 samples that are representative of central Bangladesh (Figure 2). Therefore, further surveillance might identify the extent to which AAL2 has spread and the diversity of rabies viruses in other parts of Bangladesh that might alter the estimated date of spread. It has been reported that arctic rabies virus and other variants can co-circulate in the same region (12).
The G protein is the major factor responsible for the pathogenesis of rabies virus and contains 2 glycosylation sites (13). The G protein of strains from Bangladesh uniquely evolved to contain 3 potential glycosylation sites, which has been reported in only fixed (laboratory adapted) strains and proposed to be responsible for their reduced pathogenicity (13). However, the site for additional glycosylation differs between Bangladeshi and fixed strains. Detection of an additional glycosylation site and amino acid substitutions deserve further investigations.
AAL viruses could have moved southward from Siberia or other northern regions of the former Soviet Union into Nepal, India, and other countries in Asia by a species jump from fox to dog at some point (3). Another possibility is that AAL viruses first emerged in dogs in southern Asia and subsequently spread to northern climes, where they are now maintained in fox populations (3,8). Extensive surveillance of viruses from Iran, Iraq, Afghanistan, and countries north of them is necessary to determine the origin and spread pattern of AAL rabies virus.
The timeline of divergence of different lineages determined in this study was similar to that previously reported (8). That study and our study used the full-length N gene to determine the time of divergence. Another study reported the timeline of divergence as a more recent event (14). This study used partial sequences of N genes, which might be responsible for different results. Rabies virus from Nepal also belongs to AAL2, and as reported in a previous study (15), seemed to be forming a different lineage. However, the speculation was not supported by a significant a posterior density value (0.6355). Thus, a network of countries is urgently needed to exchange information on molecular typing of circulating strains of rabies virus that might be useful in controlling rabies in this region.
Dr Jamil is a physician and virologist in the Department of Virology, Institute of Epidemiology, Disease Control, and Research, Dhaka, Bangladesh. Her research interests are virology and molecular epidemiology of rabies virus.
Acknowledgment
This study was supported in part by a Grant-in-Aid Scientific Research B from the Japan Society for the Promotion of Sciences (grant 20406026) and the Research Fund at the Discretion of the President, Oita University (grant 610000-N5010).
References
- Knobel DL, Cleaveland S, Coleman PG, Fevre EM, Meltzer MI, Miranda ME, Re-evaluating the burden of rabies in Africa and Asia. Bull World Health Organ. 2005;83:360–8. .PubMedGoogle Scholar
- Hossain M, Ahmed K, Bulbul T, Hossain S, Rahman A, Biswas MN, Human rabies in rural Bangladesh. Epidemiol Infect. 2011;140:1–8. .DOIPubMedGoogle Scholar
- Nadin-Davis SA, Turner G, Paul JP, Madhusudana SN, Wandeler AI. Emergence of Arctic-like rabies lineage in India. Emerg Infect Dis. 2007;13:111–6. .DOIPubMedGoogle Scholar
- Matsumoto T, Ahmed K, Wimalaratne O, Yamada K, Nanayakkara S, Perera D, Whole-genome analysis of a human rabies virus from Sri Lanka. Arch Virol. 2011;156:659–69. .DOIPubMedGoogle Scholar
- Drummond AJ, Rambaut A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol. 2007;7:214. .DOIPubMedGoogle Scholar
- Posada D, Crandall KA. MODELTEST: testing the model of DNA substitution. Bioinformatics. 1998;14:817–8. .DOIPubMedGoogle Scholar
- Bourhy H, Reynes JM, Dunham EJ, Dacheux L, Larrous F, Huong VT, The origin and phylogeography of dog rabies virus. J Gen Virol. 2008;89:2673–81. .DOIPubMedGoogle Scholar
- Kuzmin IV, Hughes GJ, Botvinkin AD, Gribencha SG, Rupprecht CE. Arctic and Arctic-like rabies viruses: distribution, phylogeny and evolutionary history. Epidemiol Infect. 2008;136:509–19. .DOIPubMedGoogle Scholar
- Tenzin , Dhand NK, Dorjee J, Ward MP. Re-emergence of rabies in dogs and other domestic animals in eastern Bhutan, 2005–2007. Epidemiol Infect. 2011;139:220–5. .DOIPubMedGoogle Scholar
- Nagarajan T, Nagendrakumar SB, Mohanasubramanian B, Rajalakshmi S, Hanumantha NR, Ramya R, Phylogenetic analysis of nucleoprotein gene of dog rabies virus isolates from southern India. Infect Genet Evol. 2009;9:976–82. .DOIPubMedGoogle Scholar
- Reddy GB, Singh R, Singh RP, Singh KP, Gupta PK, Desai A, Molecular characterization of Indian rabies virus isolates by partial sequencing of nucleoprotein (N) and phosphoprotein (P) genes. Virus Genes. 2011;43:13–7. .DOIPubMedGoogle Scholar
- Mansfield KL, Racloz V, McElhinney LM, Marston DA, Johnson N, Ronsholt L, Molecular epidemiological study of Arctic rabies virus isolates from Greenland and comparison with isolates from throughout the Arctic and Baltic regions. Virus Res. 2006;116:1–10. .DOIPubMedGoogle Scholar
- Yamada K, Park CH, Noguchi K, Kojima D, Kubo T, Komiya N, Serial passage of a street rabies virus in mouse neuroblastoma cells resulted in attenuation: Potential role of the additional N-glycosylation of a viral glycoprotein in the reduced pathogenicity of street rabies virus. Virus Res. 2012;165:34–45. .DOIPubMedGoogle Scholar
- Nadin-Davis SA, Sheen M, Wandeler AI. Recent emergence of the Arctic rabies virus lineage. Virus Res. 2012;163:352–62. .DOIPubMedGoogle Scholar
- Pant GR, Horton DL, Dahal M, Rai JN, Ide S, Leech S, Characterization of rabies virus from a human case in Nepal. Arch Virol. 2011;156:681–4. .DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 18, Number 12—December 2012
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Kamruddin Ahmed, Research Promotion Institute, Oita University, Yufu 879-5593, Oita, Japan
Top