Volume 18, Number 5—May 2012
Dispatch
Influenza A and B Virus Attachment to Respiratory Tract in Marine Mammals
Cite This Article
Citation for Media
Abstract
Patterns of virus attachment to the respiratory tract of 4 marine mammal species were determined for avian and human influenza viruses. Attachment of avian influenza A viruses (H4N5) and (H7N7) and human influenza B viruses to trachea and bronchi of harbor seals is consistent with reported influenza outbreaks in this species.
Understanding is limited about factors determining the ability of influenza viruses to cross the species barrier and persist in a new host population (1,2). In marine mammals, several subtypes of avian influenza A virus have caused epidemics in harbor seals (Phoca vitulina) (3–6). Also, human influenza B virus has been detected in harbor seals (7). These observations indicate the ability of both viruses to cross the species barrier and persist in harbor seals. In other marine mammal species, outbreaks of avian influenza A virus or infection with human influenza B virus have not been reported.
Attachment of influenza virus to tissues in the respiratory tract is a major determinant of host susceptibility to infection, efficiency of transmission, and pathogenicity and has been studied only to a limited degree (8,9). Attachment is determined largely by the specificity with which influenza virus attaches to sialosaccharide receptors on the host cell surface. In general, human influenza viruses prefer sialosaccharides in which sialic acid is linked to galactose by an α-2,3 linkage (SA-α-2,3-Gal), and avian influenza viruses prefer those with an α-2,6 linkage (SA-α-2,6-Gal) (10).
To understand differences in these properties between harbor seals and other marine mammals, we determined patterns of attachment for influenza virus strains known to have infected the respiratory tract of harbor seals, gray seals (Halichoerus grypus), harbor porpoises (Phocoena phocoena), and bottlenose dolphins (Tursiops truncatus). We chose gray seals, porpoises, and dolphins because their ranges overlap those of harbor seals and they are commonly kept in captivity.
We determined patterns of attachment to respiratory tract tissues of 4 sympatric marine mammal species for several influenza viruses. Avian influenza A virus subtypes H7N7 (A/Seal/Massachusetts/1/80) and H4N5 (A/Seal/Ma/47/83) were chosen because they had caused outbreaks in harbor seals (4,5). An influenza B virus strain (B/Seal/Netherlands/1/99) was chosen because it had been isolated from a harbor seal (7).
For each of these 3 viruses, we also included a closely related strain from the putative donor host species (H7N7 A/Mallard/Sweden/100/02, H4N5 A/Mallard/Netherlands/13/2008, and B/Harbin/7/94, respectively) to determine whether adaptation to the new host species was associated with a change in attachment. Influenza virus A(H1N1)pdm09 (A/Netherlands/164/09) and seasonal subtype (H3N2) virus (A/Netherlands/213/03) were chosen because they circulate endemically in humans and might have contact with captive marine mammals through their caretakers. All viruses were isolated as described (11–13).
We obtained respiratory tract specimens from marine mammals from archives of paraffin-embedded tissues. Trachea and lung (including bronchus, bronchiole, and pulmonary alveoli) from 3 animals per species were examined.
Attachment of influenza virus to tissues was visualized by histochemical analysis as described (13). A positive result by light microscopy was granular to diffuse red staining on the apical surface of epithelial cells in trachea, bronchi, and bronchioles and on alveolar cells. Staining was scored as the percentage of cells in a section showing virus attachment. We also evaluated virus attachment to submucosal glands.
Figure
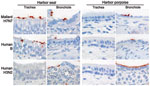
Figure. . . Attachment of 2 human influenza viruses and 1 avian influenza virus to trachea and bronchiole of harbor seal (Phoca vitulina) and harbor porpoise (Phocoena phocoena). Red staining indicates virus...
Results of attachment differed between avian influenza A viruses, human influenza A viruses, and human influenza B viruses. First, attachment of avian influenza A viruses to tracheal and bronchial epithelium was moderate in seals (harbor seal and gray seal) and absent in cetaceans (harbor porpoise and bottlenose dolphin) (Figure). Attachment to bronchiolar epithelium was moderate in seals and scarce in cetaceans, and attachment to alveolar epithelium was scarce in all 4 species. There were a few exceptions for virus attachment (Table). The source of avian influenza viruses (mallard or harbor seals) did not have a consistent effect on virus attachment in respiratory tract epithelium of any evaluated species (Table).
Second, attachment of human influenza A viruses to tracheal and bronchial epithelium was absent in seals and scarce in cetaceans. Attachment to bronchiolar and alveolar epithelium was absent or scarce in seals and moderate to abundant in cetaceans (Table, Figure). We detected few differences between attachment of influenza virus A(H1N1)pdm09 and seasonal subtype (H3N2) virus to respiratory tract tissues of cetaceans (Table).
Third, attachment of influenza B viruses to respiratory tract epithelium at all levels was scarce to moderate in seals. Attachment was negative for tracheal and bronchial epithelium, scarce for bronchiolar epithelium, and moderate for alveolar epithelium in cetaceans (Table, Figure).
Attachment of avian influenza A viruses to the respiratory tract was generally consistent with reports, or lack thereof, of avian influenza in these 4 marine mammal species. Moderate attachment of avian influenza A viruses to the trachea and bronchi of harbor seals suggests high susceptibility to and efficient transmission of these viruses. This finding is consistent with reported outbreaks of avian influenza in harbor seals (4–6). Scarce attachment of avian influenza viruses to bronchioles and alveoli of harbor seals is consistent with low pathogenicity of these viruses for harbor seals during experimental infection (4–6).
Attachment of avian influenza A virus to the respiratory tract in gray seals strongly resembles attachment in harbor seals. However, infection or outbreaks of avian influenza A virus in gray seals have not been reported, probably because virus attachment is required but is not sufficient for infection. Lack of attachment of avian influenza A viruses to trachea and bronchi of harbor porpoises and bottlenose dolphins suggests low susceptibility and inefficient transmission. This finding is consistent with lack of reported avian influenza A virus infections in these species (14).
Absence or scarcity of attachment of human influenza A viruses to trachea and bronchi of any of the marine mammal species contrasts with that of humans (13), in whom trachea and bronchi mainly express SA-α-2,6-Gal (15). This finding suggests low susceptibility to infection and can explain the lack of reported human influenza A virus infections in these 4 marine mammal species (14), even though they are often kept in captivity and are therefore at risk for infection from humans.
Attachment of influenza B virus to the respiratory tract of the 4 marine mammal species resembled that of the avian influenza A viruses. Moderate attachment of influenza B virus to the respiratory tract of seals suggests high susceptibility and efficient transmission. This finding is consistent with isolation of influenza B virus from a harbor seal and serologic evidence of influenza B virus infection in gray seals (7). Lack of attachment of influenza B virus to trachea and bronchi of cetaceans is consistent with absence of reported influenza B virus infections in these species.
Source of virus strain had little effect on its attachment. In general, there was high similarity of attachment of avian influenza A viruses from harbor seals and mallards. These findings suggest that avian influenza viruses do not require a different pattern of attachment to infect and transmit efficiently among harbor seals and that harbor seals might be susceptible to a wider range of avian influenza viruses than reported.
In conclusion, we report extensive diversity in the pattern of attachment of influenza viruses to the respiratory tract of marine mammals, which was determined by virus strain and host species involved. Our results correspond to field observations of influenza in marine mammals, i.e., outbreaks of avian influenza A virus and human influenza B virus infection in harbor seals (4,5) and lack of evidence of human influenza A virus infection in marine mammals. These results suggest that, as in humans (11,15), attachment of influenza virus to the proximal part of the respiratory tract, which depends largely on appropriate sialic acid moieties, is critical for susceptibility and efficient transmission of influenza viruses in marine mammals.
Dr Ramis is an associate professor of veterinary pathology at the Universitat Autònoma de Barcelona, Spain. His research interests are avian and wildlife pathology and infectious diseases.
Acknowledgments
We thank Yoshihiro Kawaoka and Martha McGregor for providing the seal (H4N5) virus (A/Seal/Ma/47/83), Michelle Fleetwood for providing some of the Tursiops truncatus tissues, and F. van der Panne and P. van Run for technical assistance.
A.J.R. was supported by the government of Spain (PR2009-0145).
References
- Olsen B, Munster VJ, Wallensten A, Walsdenström J, Osterhaus AD, Fouchier RA. Global patterns of influenza A virus in wild birds. Science. 2006;312:384–8. DOIPubMedGoogle Scholar
- Lamb RA, Krug RM. Orthomyxoviridae: the viruses and their replication. In: Knipe DM, Howley PM editors. Fields virology. 4th ed. Philadelphia: Lippincot Williams and Wilkins; 2001. p. 1487–1531.
- Callan RJ, Early G, Kida H, Hinshaw VS. The appearance of H3 influenza viruses in seals. J Gen Virol. 1995;76:199–203. DOIPubMedGoogle Scholar
- Geraci JR, St Aubin DJ, Barker IK, Webster RJ, Hinshaw VS, Bean WJ, Mass mortality of harbor seals: pneumonia associated with influenza A virus. Science. 1982;215:1129–31. DOIPubMedGoogle Scholar
- Hinshaw VS, Bean WJ, Webster RJ, Rehg JE, Fiorelli P, Early G, Are seals frequently infected with avian influenza viruses? J Virol. 1984;51:863–5.PubMedGoogle Scholar
- Webster RG, Hinshaw VS, Bean WJ, Van Wyke KL, Geraci JR, St Aubin DJ. Characterization of an influenza A virus from seals. Virology. 1981;113:712–24. DOIPubMedGoogle Scholar
- Osterhaus AD, Rimmelzwaan GF, Martina BE, Bestebroer TM, Fouchier RA. Influenza B virus in seals. Science. 2000;288:1051–3. DOIPubMedGoogle Scholar
- Kuiken T, Holmes EC, McCauley J, Rimmelzwaan GF, Williams CS, Grenfell BT. Host species barriers to influenza virus infections. Science. 2006;312:394–7. DOIPubMedGoogle Scholar
- Ito T, Kawaoka Y, Nomura A, Otsuki K. Receptor specificity of influenza A viruses from sea mammals correlates with lung sialyloligosaccharides in these animals. J Vet Med Sci. 1999;61:955–8. DOIPubMedGoogle Scholar
- Rogers GN, Paulson JC. Receptor determinants of human and animal influenza virus isolates: differences in receptor specificity of the H3 hemagglutinin based on species origin. Virology. 1983;127:361–73. DOIPubMedGoogle Scholar
- van Riel D, de Bakker MA, Leijten L, Chutinimitkul S, Munster V, de Wit E, Seasonal and pandemic human influenza viruses attach better to human upper respiratory tract epithelium than avian influenza viruses. Am J Pathol. 2010;176:1614–8. DOIPubMedGoogle Scholar
- van Riel D, Muster V, de Wit E, Rimmelzwaan GF, Fouchier RA, Osterhaus AD, H5N1 virus attachment to lower respiratory tract. Science. 2006;312:399. DOIPubMedGoogle Scholar
- van Riel D, Muster V, de Wit E, Rimmelzwaan GF, Fouchier RA, Osterhaus AD, Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am J Pathol. 2007;171:1215–23. DOIPubMedGoogle Scholar
- Reperant LA, Rimmelzwaan GF, Kuiken T. Avian influenza viruses in mammals. Rev Sci Tech. 2009;28:137–59.PubMedGoogle Scholar
- Shinya K, Ebina M, Yamada S, Ono M, Kasai N, Kawaoka Y. Avian flu: influenza virus receptors in the human airway. Nature. 2006;440:435–6. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleTable of Contents – Volume 18, Number 5—May 2012
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Antonio J. Ramis, Facultat de Veterinària, Universitat Autònoma de Barcelona, 08193-Bellaterra, Barcelona, Spain
Top