Volume 17, Number 7—July 2011
Research
Co-infections of Plasmodium knowlesi, P. falciparum, and P. vivax among Humans and Anopheles dirus Mosquitoes, Southern Vietnam
Cite This Article
Citation for Media
Abstract
A single Anopheles dirus mosquito carrying sporozoites of Plasmodium knowlesi, P. falciparum, and P. vivax was recently discovered in Khanh Phu, southern Vietnam. Further sampling of humans and mosquitoes in this area during 2009–2010 showed P. knowlesi infections in 32 (26%) persons with malaria (n = 125) and in 31 (43%) sporozoite-positive An. dirus mosquitoes (n = 73). Co-infections of P. knowlesi and P. vivax were predominant in mosquitoes and humans, while single P. knowlesi infections were found only in mosquitoes. P. knowlesi–co-infected patients were largely asymptomatic and were concentrated among ethnic minority families who commonly spend nights in the forest. P. knowlesi carriers were significantly younger than those infected with other malaria parasite species. These results imply that even if human malaria could be eliminated, forests that harbor An. dirus mosquitoes and macaque monkeys will remain a reservoir for the zoonotic transmission of P. knowlesi.
Concerted control measures have considerably reduced the prevalence of malaria in Vietnam, and the parasites that cause it are now mostly restricted to forested rural areas (1). Forest malaria poses a special challenge for control because the exophilic and early biting habits of the mosquito vector Anopheles dirus render conventional vector control methods such as indoor residual spraying and insecticide treated mosquito nets difficult to apply as well as ineffective (2–4). The possibility of zoonotic malaria in Southeast Asian forests, because of the transmission of Plasmodium knowlesi from monkeys to humans (5–10), may form an additional complication.
Since surveys began in 2002, the forest populations of An. dirus mosquitoes in Khanh Phu, south-central Vietnam, have shown consistently high sporozoite infection rates (1%–2% of the thousands of specimens collected and dissected annually for microscopic examination of salivary glands), raising the question of whether all the sporozoites detected belong to species capable of infecting humans. In 2008, evidence was found for the co-infection of P. knowlesi, P. falciparum, and P. vivax in the salivary glands of 1 mosquito among 17 that had been processed by PCR with malaria parasite species–specific primers (11). Here we report the results of the PCR analysis of 72 additional sporozoite–positive salivary glands of An. dirus mosquitoes from the forest in Khanh Phu and of 211 blood samples from the local human population.
Study Area
Khanh Phu (12°14′N; 108°56′E) is a commune with ≈3,000 inhabitants, mainly of the Raglai ethnic minority, who live between the forested foothills on the east side of the Truong Son mountain range in south-central Vietnam (Khanh Hoa Province), an area where malaria was previously hyper- to holo-endemic (3). Since 1993, the Medical Committee Netherlands–Vietnam (a Dutch nongovernmental organization) has cooperated with the National Institute of Malariology, Parasitology and Entomology in Hanoi; the Institute of Malariology, Parasitology and Entomology in Qui Nhon; and the Malaria Control Centre of Khanh Hoa Province to set up and operate the Khanh Phu Malaria Field Research Unit to provide Vietnamese malaria researchers with the opportunity to study local malaria epidemiology and develop and test improved control methods. An. dirus species A is currently the only malaria-transmitting species of mosquitoes in Khanh Phu. Anopheles minimus mosquitoes, previously the major malaria vector in the region, disappeared from Khanh Phu after 1998, following the wide-scale use of insecticide-treated mosquito nets (3,12). While the average malaria prevalence in the human population has been greatly reduced (from >50% before 1998 to 2%–3% during 2003–2009), persons who sleep overnight in the forest still run a high risk for infection. Analysis of samples from infected persons by thin-smear microscopy showed that approximately two thirds were caused by P. falciparum, one third by P. vivax, and a very small number by P. malariae during the 2003–2009 study period. This malaria prevalence typically affects the poorest members of the local Raglai ethnic community, whose livelihood partly depends on excursions into the forest to collect products such as bamboo and rattan, or to cultivate their plots on the mountain slopes. All residences within the commune were mapped, and all persons were registered at the research station and assigned unique code numbers based on residential location and family relations. (All methods in this study that involved human participants in the field were certified as permitted standard procedures by the National Institute of Malariology, Parasitology and Entomology in Hanoi.)
Mosquito Collection and Salivary Gland Examination
Mosquitoes were collected by human-baited landing catches at 4 collection sites in the forest and forest fringe areas near Nga Hai village in the southern part of Khanh Phu commune, from January 2008 through February 2010. Mosquito collectors were adult men of the Raglai ethnic group. They were intensively screened for malaria and promptly treated with artemisinin combination therapy if infected. The collectors worked in teams of 2 over the whole night, 1 person collected from 6:00
Anopheles species were determined on the basis of morphologic features (13). All An. dirus group mosquitoes were assumed to be An. dirus species A on the basis of previous accurate identifications and the known distribution of this species (14,15). Female anopheline mosquitoes were dissected for salivary glands, midgets, and ovaries, and these were examined by microscopy for the presence of sporozoites, oocysts, and parity, respectively. Sporozoite-infected glands were applied to filter paper and dried in an ambient atmosphere before storage in closed vials at 4°C–6°C.
Collection of Human Blood Samples
Blood samples were collected by 2 methods. First, to detect as many parasite-infected persons as possible, from March 8, 2009, through February 28, 2010, blood was collected by targeted active case detection from persons who had frequently worked in the forest or had reported fever. A total of 549 blood samples from 305 persons from 156 families were collected; 183 persons were sampled once, and no one was sampled >8 times; the sex ratio had a male bias (64% male). Sixty-nine of the 121 blood samples that were positive by microscopy, and an additional randomly selected 105 of the negative samples were processed by PCR to determine Plasmodium species. Second, from March 11, 2010, through March 30, 2010, a cross-sectional survey was undertaken in the 2 villages nearest to the mosquito collection sites. A single blood sample was taken from each of 624 residents, irrespective of symptoms or work place. One hundred and thirty-five persons in this sample had previously given samples as part of the targeted active case detection (ACD) survey; 489 had not given samples previously. From the 49 blood samples found positive by microscopy, 37 were randomly chosen for PCR processing.
Sixty-eight percent of all blood slides in both samples were from persons in the 2 southernmost villages of Khanh Phu (Nga Hai and Da Trai). This represents an area where ≈1,100 persons are living, spread over an area of 1.7 km2 at 12°12.5′N and 108°55.5′E. All their houses are located <1 km from the nearest forest and 1–3 km away from the mosquito collection sites. An. dirus mosquitoes were rarely caught in these villages.
Blood was collected by finger-prick; thick and thin blood films were made for diagnosis, and blood was applied to filter paper for downstream molecular analyses. All adult volunteers provided informed consent and for children, consent was obtained from close relatives. All persons found to be infected with parasites (as diagnosed by microscopy) received treatment, according to the policy of the Vietnam Ministry of Health.
DNA Extraction and Parasite Species Identification by PCR
DNA was extracted from dried blood samples on filter paper and from sporozoite-infected glands, and subsequent malaria parasite species identification by PCR that targeted the 18S rRNA gene was carried out as previously described (11,16). Briefly, DNA was extracted by using the QIAamp DNA micro kit (QIAGEN, Tokyo, Japan). Extracted DNA samples were stored at –20°C until use. Plasmodium species–specific nested PCR assays to detect and identify human malaria parasites were performed as described (17). For detection of the P. knowlesi 18S rRNA gene, the primers Pmk8 and Pmk9 were used (5). Because this primer set can occasionally cross-react with P. vivax DNA and produce false-positive results (18), samples were also subject to a further PCR targeting of the P. knowlesi circumsporozoite protein (CSP) gene as described (8). PCR products were separated by electrophoresis on 1.5% agarose gels and stained with ethidium bromide. Primer sequences for the gene of human Plasmodium species (17), 18S rRNA of P. knowlesi (5), and the CSP gene of P. knowlesi (8) were as previously described. P. knowlesi H strain (American Type Culture Collection no. 30158) (kindly donated by Satoru Kawai, Dokkyou University, Japan) was used as a positive control. We verified that no cross-reaction occurred between the primer sets used to amplify P. vivax CSP and P. knowlesi CSP by using DNA extracted from single infections of both species (data not shown).
Statistical Analysis
Student 1- or 2-tailed t tests were used for comparing means. Chi-square tests for association and confidence limits were calculated with Microsoft Excel (Microsoft Corp., Redmond, WA, USA).
Plasmodium Infections of An. dirus Mosquitoes
From January 1, 2008, through February 22, 2010, a total of 6,834 female anopheline mosquitoes were captured, of which 83.2% belonged to An. dirus s.s., and 8.6% to An. maculatus, 5.4% to An. peditaeniatus; the remaining 2.8% were divided between a further 11 anopheline species in very small numbers. Of the 8,331 An. maculatus mosquitoes dissected in Khanh Phu over >15 years, only 1 mosquito was ever found to be infected with sporozoites. None of the other Anopheles species, with the exception of An. dirus, were ever found to be infected with sporozoites. Thus An. dirus mosquitoes are regarded as the only malaria vectors in the Khanh Phu forest.
An. dirus mosquitoes were found during every month of sampling with an average human-biting density of 4.4 bites/person-night (range 0.3–17.4; Table A1). As in previous years in Khanh Phu (3), these mosquitoes usually reached peak densities in the dry season (February–April). Numbers were lowest during the hottest part of the year (May–August), climbed to a second peak with the onset of the rains during September–November, and usually decline again following heavy rains during October–December. The monthly average parous rate fluctuated between 65% and 90% (average 77%).
Sporozoites were detected by microscopic screening in 89 (1.57%) of 5,663 dissected An. dirus mosquitoes, and oocysts were observed on 0.94% of the midguts. The sporozoite rate showed some seasonal fluctuations, but these did not clearly correlate with the fluctuations in biting density. Multiplying the sporozoite rate by the human-biting rate gave an average annual entomological inoculation rate of 25.4 infective bites per person per year. This is comparable with the average rate estimated during the previous 8 years in the forest of Khanh Phu (22 infective bites per person per year, unpub. data). However, both the sporozoite rate (4.6%) and human-biting density (12 bites/person-night) reached a maximum in the last 2 months of the survey (January and February 2010). This finding implies that persons who slept unprotected in the forest during this period would have had a >50% chance of being bitten by an infected An. dirus mosquito during any 1 night.
Figure 1

Figure 1. Results and interpretation of the PCR analyses of sporozoite-positive salivary glands of Anopheles dirus mosquitoes in Khanh Phu forest, Vietnam. CSP, circumsporozoite protein; ssu, small subunit; shaded cells, PCR products present;...
Of the 86 sporozoite-positive mosquitoes, 73 underwent PCR analysis for malaria parasite detection, of which 72 were successfully assayed. Thirty-one (43%) of these 72 salivary glands were PCR positive for P. knowlesi csp and for P. knowlesi 18S rRNA. Five additional specimens were positive for P. knowlesi 18S rRNA but negative for P. knowlesi CSP and were therefore regarded as P. knowlesi negative (Figure 1). One specimen was only positive for P. knowlesi CSP and not by any other test, and therefore the species could not be confidently determined. The frequencies of P. falciparum, P. vivax, and P. malariae were: 50%, 50%, and 6%, respectively. In 22 (71%) of 31 glands, P. knowlesi was found as a co-infection with one of these other species. The combination P. knowlesi + P. vivax (in 14 glands) was far more common than P. knowlesi + P. falciparum (1 case). The combination of P. knowlesi + P. vivax + P. falciparum was, however, also quite common (7 glands).
Figure 2

Figure 2. Dynamics of biting density and sporozoite positive salivary glands (including Plasmodium knowlesi) of Anopheles dirus mosquitoes in Khanh Phu forest, Vietnam. The solid red line connects the points of mean monthly...
The frequency of P. knowlesi–positive mosquitoes did not significantly differ between the 4 collections sites, and P. knowlesi was detected during 8 months of the past year, except in August, September, November, and December when sporozoite-infected mosquitoes were few (Figure 2). However, the highest number of sporozoite-infected mosquitoes and 13 of the 31 P. knowlesi infections were found in January 2010.
Plasmodium Infections of Humans
Of the 549 blood samples collected by targeted ACD from March 2009 through February 2010, 121 (22.0%) were parasite positive by microscopy, including 94 (17.1%) that were positive for P. falciparum. The cross-sectional survey conducted in March 2010 showed a parasite prevalence of 7.9% (10.5% in male residents and 5.2% in female residents; χ2 p = 0.013). The P. falciparum prevalence was 5.1% (7.3% in male residents and 2.9% in female residents; χ2 p = 0.012).
The Table shows the results of parasite species identification by microscopy and PCR for both the targeted ACD and cross-sectional surveys. Both samples had a male bias. In the targeted ACD sample, this result reflected the skewed sex ratio in the total 549 blood samples. Among the 624 persons sampled during the cross-sectional survey the sex ratio was 50/50, but because infections were more common in men, the random subsample of 37 for PCR analysis of the 49 positive results by microscopy was also male biased. The mean age of women and men was similar, and the difference in mean age between findings of the targeted ACD and those of the cross-sectional survey was not significant (t test p>0.05). In the targeted ACD sample, 19 (18%) of 105 samples that were negative by microscopic examination were parasite positive by PCR. Five patients with positive results in the ACD were shown by PCR to have P. malariae as a co-infection, and 1 patient in the cross-sectional survey had a P. malariae single infection, all of which were not seen by microscopy. We were unable to determine the parasite densities of P. knowlesi in our samples because it was not possible to discriminate between parasite species in co-infections at low parasite densities.
A total of 32 blood samples were found to be infected with P. knowlesi on the basis of simultaneous positivity in the P. knowlesi 18S rRNA and P. knowlesi csp PCR tests: 19 (51%) of 37 malaria-infected persons in the cross-sectional survey, and 13 (15%) of 88 PCR-positive cases collected by targeted ACD. Four additional specimens in the ACD sample were positive for P. knowlesi 18S rRNA but negative for P. knowlesi csp, all in co-infections with other parasite species, most commonly P. vivax. Human blood samples positive for P. knowlesi csp and negative for P. knowlesi 18S rRNA were not found.
P. knowlesi infection was detected by targeted ACD in 8 of 12 months. On the basis of the PCR results of the samples from the ACD survey, the annual P. knowlesi incidence can be estimated at 10/1,000 person-years (95% confidence limit 4%–17%). The results from the cross-sectional survey imply a peak P. knowlesi prevalence in March 2010 among the population of the southern villages of 12.6% (95% confidence limit 6.3%–21.1%). The large increase in P. knowlesi co-infections in humans in March 2010 correlates with the increased frequency of this parasite in the mosquitoes in the preceding months.
Twenty-eight of the 32 P. knowlesi infections were found in persons of Raglai ethnicity, the dominant group in the study area. However, 2 of the 5 persons of Trinh ethnicity and 2 of the 7 persons of Kinh ethnicity sampled also had a P. knowlesi co-infection; all had asymptomatic cases detected during the cross-sectional survey.
Figure 3
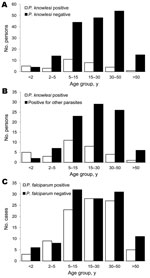
Figure 3. Age analysis of persons tested for Plasmodium knowlesi infection, Khanh Phu, Vietnam. A) Age groups of P. knowlesi–positive persons (n = 32; mean age 15.8 y) compared with P. knowlesi–negative persons...
The average age of persons with P. knowlesi co-infections was 15.0 years in the targeted ACD survey and 16.9 years in the cross-sectional survey; both ages were significantly lower than the ages of P. knowlesi–infected persons in both samples (24.8 and 26.2 years, respectively; 2-tailed t test p<0.001; Figure 3, panel A) and those of all other positive case-patients (24.8 and 24.1 years, respectively; 2-tailed t test p< 0.01; Figure 3, panel B). P. falciparum–infected persons had a mean age of 23.4 years, which did not differ significantly from the mean age of 25.6 years for P. falciparum–negative persons (2-tailed t test p>0.3; Figure 3, panel C).
Six (19%) of the 32 P. knowlesi co-infected persons had fever. This finding was not significantly different from the 10% with fever among 86 uninfected persons in the PCR-analyzed sample (χ2 p = 0.23). Only persons with single P. falciparum infections were significantly more often febrile (30%) than uninfected persons (χ2 p = 0.003).
Figure 4
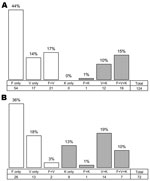
Figure 4. Frequency of single and co-infections among 124 human blood samples (A) and 73 mosquito salivary glands (B) positive for Plasmodium spp. infection by PCR. P. malariae was discarded. Gray bars indicate...
In the human blood samples, P. knowlesi was only ever found in a co-infection, almost always with P. vivax and often with P. falciparum in addition. The lack of the P. knowlesi and P. falciparum without P. vivax combination (only 1 case each in the mosquito and human samples; Figure 4, panels A and B) contributed most strongly to the significance of the nonrandom distribution (χ2 p<0.001). P. knowlesi infection was equally frequent among PCR-positive women and men, whereas P. falciparum was significantly more frequent among the PCR-positive men (Table).
P. knowlesi occurs frequently in humans in Khanh Phu as well as in the An. dirus mosquito population in nearby forests. This finding, in combination with the increasing number of reports of P. knowlesi infections from Thailand (6) and Myanmar (10), in similar forested environments, highlights the wide range of this pathogen in humans in Southeast Asia.
Our results provide additional information about vector bionomics and the clinical manifestation and epidemiology of P. knowlesi in a native population. P. knowlesi occurs in high frequencies as co-infections in An. dirus species A mosquitoes, which is the only human malaria vector in this area (3). Other anopheline species thus far proven to transmit P. knowlesi between macaque monkeys and humans in Southeast Asia, A. cracens and A. latens (8,19), belong, together with An. dirus species, to the Leucosphyrus group. Due to the widespread distribution of several An. dirus species in Southeast Asia (14) a prominent role of An. dirus sensu lato in sylvatic and zoonotic malaria transmission seems likely.
In our study, P. knowlesi infections in humans were always associated with infections of other Plasmodium species. This finding was in contrast to infections in mosquitoes, in which P. knowlesi single infections were relatively common. Furthermore, the combination of P. falciparum + P. knowlesi was far less common in both humans and mosquitoes than would be expected by chance. This may suggest a degree of interaction between these species that precludes the establishment of co-infections. Such nonrandom association was, however, not found in studies from Thailand, Malaysia, and Myanmar (6,8,10).
Even though known to be potentially dangerous for humans (20–22), the P. knowlesi co-infections described here did not lead to severe disease or fever and were concentrated in a group of ethnic minority families who often work in or near the forest, confirming a report from a neighboring area (23). The lack of symptoms, high sensitivity of the PCR method, and the fact that neither P. knowlesi nor P. malariae (which is often mistaken for P. knowlesi) was observed by microscopy suggest that the human P. knowlesi co-infections in this area cause very low levels of parasitemia. P. knowlesi–infected persons were on average significantly younger (15.8 years) than those infected with other species (23.9 years), which suggests that natural immunity is more easily acquired against P. knowlesi than against P. falciparum and P. vivax.
Macaques are common in the forests of Khanh Phu and are likely to be bitten by the same An. dirus population that bites humans. Whether these monkeys harbor a zoonotic malaria reservoir of P. knowlesi or if the parasite is also transmitted from person to person is currently unknown and requires further investigation. In this study, only P. knowlesi, P. falciparum, P. vivax, and P. malariae were investigated. Therefore whether P. knowlesi is the only malaria parasite in monkeys transmitted by An. dirus mosquitoes in this area is unknown.
Recent discoveries have demonstrated the relative ease and frequency at which malaria parasites may have switched between hosts (24–29). The factors that influence the probability of such host switching are likely to be many and varied, but the presence of a mosquito vector that brings the parasites in contact with different hosts must be a key precondition. An. dirus species A appears to fit this role.
Epidemiologic and Public Health Implications
These findings may fundamentally change the perspectives for the control of forest malaria. Previously, forest malaria may have been considered manageable because the parasite reservoir in the forest may be reduced through intensive case detection and treatment of human communities living in or near the forests. The likely presence of a nonhuman reservoir of P. knowlesi (and possibly other parasites) in monkeys and of a mosquito vector that intensively inoculates parasites among monkeys and humans reinforces the need to find methods of vector control or biting prevention that can be applied to An. dirus mosquitoes.
Mr Marchand has been a senior advisor for the Medical Committee Netherlands–Vietnam for 20 years. He has developed and guided various projects in malaria and dengue operational research as well as in rural development among poor mountainous communities in Vietnam.
Acknowledgments
We thank Nguyen Son Hai, Phan Chau Do, Dang Duy Vu and Nguyen Le Dung for their help in collecting and processing the mosquito and blood samples; the mosquito collectors for their hard work and long nights in the forest; and the residents of Khanh Phu for their willingness to contribute blood samples over many years. We also thank the Provincial Health Service and Malaria Control Centre of Khanh Hoa Province as well as the authorities of Khanh Vinh district and Khanh Phu municipality for their administrative support.
The Khanh Malaria Research team and the field research costs were financially supported through private donations to the Medical Committee Netherlands–Vietnam. The laboratory work in Japan was partly supported by a grant-in-aid for the Japanese-Vietnam Collaborative Research Program from the Japan Society for the Promotion of Science and a grant-in-aid for Scientific Research from Fujita Health University.
References
- Viet Nam National Institute of Malariology. Parasitology and Entomology (NIMPE). Annual reports of the National Malaria Control Program in Vietnam: 2003–2008. Hanoi: The Institute; 2008.
- Erhart A, Thang ND, Hung NQ, Toi LV, Hung LX, Tuy TQ, Forest malaria in Vietnam: a challenge for control. Am J Trop Med Hyg. 2004;70:110–8.PubMedGoogle Scholar
- Marchand RP. The Khanh Phu malaria research project: an overview 1994–2004. 2005. [cited 2010 Sep 4]. http://www.mcnv.nl/uploads/media/Malaria_overview_2005_18.pdf
- Erhart A, Ngo DT, Phan VK, Ta TT, Van Overmeir C, Speybroeck N, Epidemiology of forest malaria in central Vietnam: a large scale cross-sectional survey. Malar J. 2005;4:58. DOIPubMedGoogle Scholar
- Singh B, Kim Sung L, Matusop A, Radhakrishnan A, Shamsul SS, Cox-Singh J, A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet. 2004;363:1017–24. DOIPubMedGoogle Scholar
- Putaporntip C, Hongsrimuang T, Seethamchai S, Kobasa T, Limkittikul K, Cui L, Differential prevalence of Plasmodium infections and cryptic Plasmodium knowlesi malaria in humans in Thailand. J Infect Dis. 2009;199:1143–50. DOIPubMedGoogle Scholar
- Luchavez J, Espino F, Curameng P, Espina R, Bell D, Chiodini P, Human infections with Plasmodium knowlesi, the Philippines. Emerg Infect Dis. 2008;14:811–3. DOIPubMedGoogle Scholar
- Vythilingam I, Noorazian YM, Huat TC, Jiram AI, Yusri YM, Azahari AH, Plasmodium knowlesi in humans, macaques and mosquitoes in peninsular Malaysia. Parasit Vectors. 2008;1:26. DOIPubMedGoogle Scholar
- Cox-Singh J, Singh B. Knowlesi malaria: newly emergent and of public health importance? Trends Parasitol. 2008;24:406–10. DOIPubMedGoogle Scholar
- Jiang N, Chang Q, Sun X, Lu H, Yin J, Zhang Z, Co-infections with Plasmodium knowlesi and other malaria parasites, Myanmar. Emerg Infect Dis. 2010;16:1476–8. DOIPubMedGoogle Scholar
- Nakazawa S, Marchand RP, Quang NT, Culleton R, Manh ND, Maeno Y. Anopheles dirus co-infection with human and monkey malaria parasites in Vietnam. Int J Parasitol. 2009;39:1533–7. DOIPubMedGoogle Scholar
- Garros C, Marchand RP, Quang NT, Hai NS, Manguin S. First record of Anopheles minimus C and significant decrease of An. minimus A in central Vietnam. J Am Mosq Control Assoc. 2005;21:139–43. DOIPubMedGoogle Scholar
- Vietnam National Institute of Malariology. Parasitology and Entomology. Identification key for Anophelinae in Vietnam. Hanoi (Vietnam): Medical Publishing House; 2008.
- Trung HD, Van Bortel W, Sochantha T, Keokenchanh K, Quang NT, Cong LD, Malaria transmission and major malaria vectors in different geographical areas of Southeast Asia. Trop Med Int Health. 2004;9:230–7. DOIPubMedGoogle Scholar
- Obsomer V, Defourny P, Coosemans M. The Anopheles dirus complex: spatial distribution and environmental drivers. Malar J. 2007;6:26. DOIPubMedGoogle Scholar
- Maeno Y, Nakazawa S, Dao le D, Yamamoto N, Giang ND, Van Hanh T, . A dried blood sample on filter paper is suitable for detecting Plasmodium falciparum gametocytes by reverse transcription polymerase chain reaction. Acta Trop. 2008;107:121–7. DOIPubMedGoogle Scholar
- Singh B, Bobogare A, Cox-Singh J, Snounou G, Abdullah MS, Rahman HA. A genus- and species-specific nested polymerase chain reaction malaria detection assay for epidemiologic studies. Am J Trop Med Hyg. 1999;60:687–92.PubMedGoogle Scholar
- Imwong M, Tanomsing N, Pukrittayakamee S, Day NP, White NJ, Snounou G. Spurious amplification of a Plasmodium vivax small-subunit RNA gene by use of primers currently used to detect P. knowlesi. J Clin Microbiol. 2009;47:4173–5. DOIPubMedGoogle Scholar
- Tan CH, Vythilingam I, Matusop A, Chan ST, Singh B. Bionomics of Anopheles latens in Kapit, Sarawak, Malaysian Borneo in relation to the transmission of zoonotic simian malaria parasite Plasmodium knowlesi. Malar J. 2008;7:52. DOIPubMedGoogle Scholar
- Jongwutiwes S, Putaporntip C, Iwasaki T, Sata T, Kanbara H. Naturally acquired Plasmodium knowlesi malaria in human, Thailand. Emerg Infect Dis. 2004;10:2211–3.PubMedGoogle Scholar
- Daneshvar C, Davis TME, Cox-Singh J, Rafe’ee MZ, Zakaria SK, Divis PCS, Clinical and laboratory features of human Plasmodium knowlesi infection. Clin Infect Dis. 2009;49:852–60. DOIPubMedGoogle Scholar
- Cox-Singh J, Hiu J, Lucas SB, Divis PC, Zulkarnaen M, Chandran P, Severe malaria—a case of fatal Plasmodium knowlesi infection with post-mortem findings: a case report. Malar J. 2010;9:10. DOIPubMedGoogle Scholar
- Van den Eede P, Van HN, Van Overmeir C, Vythilingam I, Duc TN, Hung LX, Human Plasmodium knowlesi infections in young children in central Vietnam. Malar J. 2009;8:249. DOIPubMedGoogle Scholar
- Escalante AA, Cornejo OE, Freeland DE, Poe AC, Durrego E, Collins WE, A monkey's tale: the origin of Plasmodium vivax as a human malaria parasite. Proc Natl Acad Sci U S A. 2005;102:1980–5. DOIPubMedGoogle Scholar
- Hayakawa T, Culleton R, Otani H, Horii T, Tanabe K. Big bang in the evolution of extant malaria parasites. Mol Biol Evol. 2008;25:2233–9. DOIPubMedGoogle Scholar
- Duarte AM, Malafronte RS, Cerutti C Jr, Curado I, de Paiva BR, Maeda AY, Natural Plasmodium infections in Brazilian wild monkeys: reservoirs for human infections? Acta Trop. 2008;107:179–85. DOIPubMedGoogle Scholar
- Prugnolle F, Durand P, Neel C, Ollomo B, Ayala FJ, Arnathau C, African great apes are natural hosts of multiple related malaria species, including Plasmodium falciparum. Proc Natl Acad Sci U S A. 2010;107:1458–63. DOIPubMedGoogle Scholar
- Garamszegi LZ. Patterns of co-speciation and host switching in primate malaria parasites. Malar J. 2009;8:110. DOIPubMedGoogle Scholar
- Liu W, Li Y, Learn GH, Rudicell RS, Robertson JD, Keele BF, Origin of the human malaria parasite Plasmodium falciparum in gorillas. Nature. 2010;467:420–5. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 17, Number 7—July 2011
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Ron P. Marchand, Khanh Phu Malaria Research Unit, MCNV Office in Khanh Hoa Province, GPO Box 19, Nha Trang City, Vietnam
Top