Volume 32, Number 6—June 2026
Synopsis
Emergence of Klebsiella pneumoniae Carbapenemase–Producing K. pneumoniae with Penicillin-Binding Protein 3 Insertions, Taiwan, 2021
Cite This Article
Citation for Media
Abstract
Carbapenem-resistant Enterobacterales (CRE) are a major global health threat with limited treatment options. Aztreonam/avibactam is a promising therapy against metallo-β-lactamase (MBL)–producing and other CRE, but emerging resistance threatens its effectiveness. Insertions in penicillin-binding protein 3 (PBP3), which are well described in Escherichia coli, are linked to reduced aztreonam/avibactam susceptibility but remain poorly characterized in Klebsiella pneumoniae. We report clinical K. pneumoniae carbapenemase–producing K. pneumoniae sequence type 11 isolates carrying a novel PBP3 YRIT insertion, conferring reduced susceptibility to aztreonam/avibactam and ceftazidime/avibactam. Functional and genetic studies suggest that the PBP3 insertion impairs β-lactam binding and, in combination with blaKPC-2 and other β-lactamases, contributes to reduced susceptibility. Those findings demonstrate the emergence of a PBP3 insertion in a high-risk K. pneumoniae clone, underscoring the expansion of this resistance mechanism and the critical need for genomic surveillance and novel therapeutics to identify and treat such infections.
Carbapenem-resistant Enterobacterales (CRE) have emerged as one of the most urgent antimicrobial resistance threats worldwide (1). Infections caused by CRE are associated with prolonged hospitalization, limited therapeutic options, and substantial mortality rates. Among CRE, strains producing metallo-β-lactamase (MBL) are of particular concern because MBL enzymes hydrolyze most β-lactams (except for aztreonam), including carbapenems, and are not inhibited by available β-lactamase inhibitors, such as clavulanate, tazobactam, avibactam, relebactam, and vaborbactam. The global spread of MBL-producing Enterobacterales, particularly New Dehli metallo-β-lactamase (NDM)–producing strains, poses a formidable challenge for clinicians and public health systems. Of note, a recent study documented a 461% increase in the age-adjusted incidence of NDM-producing CRE in the United States during 2019–2023 (2).
Aztreonam/avibactam is a promising therapeutic option for infections caused by MBL-producing organisms (3). Aztreonam is intrinsically stable to hydrolysis by MBLs, and avibactam provides inhibition to coproduced serine β-lactamases, such as extended-spectrum β-lactamase (ESBL) and Klebsiella pneumoniae carbapenemase (KPC). The aztreonam/avibactam drug combination overcomes common combined resistance mechanisms in MBL-producers and other CRE, addressing a major therapeutic gap.
Nevertheless, emerging resistance mechanisms threaten the durability of aztreonam/avibactam. One concerning development is the insertion of motifs containing 4 amino acids, most commonly YRIK or YRIN (4), into penicillin-binding protein 3 (PBP3). PBP3 is a target of aztreonam and other β-lactam antibiotics, including ceftazidime, cefepime, and cefiderocol (4–7). Those PBP3 insertions are thought to alter access to the transpeptidase pocket, thereby reducing the activity of PBP3-targeting agents. On their own, PBP3 insertions confer only modest increases in MICs to aztreonam/avibactam and ceftazidime/avibactam (5,6,8,9). However, those mutations frequently occur alongside additional β-lactamases (e.g., blaNDM and blaCMY) and other resistance determinants (e.g., cirA mutation) (10–12). Combined, those mechanisms can drive near pan–β-lactam resistance, which can have serious clinical consequences, including treatment failure and patient death (11,12).
PBP3 insertions predominantly have been described in Escherichia coli (4); however, the occurrence and clinical significance of PBP3 insertions in K. pneumoniae, another multidrug-resistant pathogen and leading cause of healthcare-associated infections, remain poorly understood. A few amino acid substitutions in PBP3 of K. pneumoniae have been implicated in increased resistance to aztreonam/avibactam, ceftazidime/avibactam, or ceftibuten/avibactam (13–15). However, to our knowledge, PBP3 insertions in clinical cases of K. pneumoniae have not yet been described in the literature, and the interplay between the PBP3 alterations and β-lactamases warrants further investigation. In this study, we describe 2 clinical cases in Taipei, Taiwan, involving KPC-producing K. pneumoniae isolates carrying PBP3 insertions.
Case 1
The first case occurred in a woman in her mid-70s with a history of heart failure and hypertension who was admitted to a tertiary-care hospital in Taipei for gastrointestinal bleeding in mid-April 2021. She initially received ciprofloxacin for pyuria on the basis of a prior urine culture that had yielded E. coli. During that admission, she had acute osteomyelitis of the right great toe diagnosed and underwent sequestrectomy on hospitalization day 14. Cultures from that specimen grew Enterococcus faecalis and Streptococcus constellatus, and she was treated with moxifloxacin. On hospitalization day 24, a new urinary tract infection developed, and urine culture grew carbapenem-resistant K. pneumoniae (isolate no. LC1490). Ceftazidime/avibactam therapy was initiated but was switched to amikacin after susceptibility testing confirmed reduced susceptibility to ceftazidime/avibactam (MIC 8 µg/mL) (Table). After a 7-day course of amikacin, the patient recovered and was discharged. In mid-to-late June 2022, she was readmitted for a right intertrochanteric hip fracture and underwent open reduction and internal fixation. Postoperatively, respiratory failure and shock developed, requiring admission to the intensive care unit. She was treated sequentially with piperacillin/tazobactam and ceftriaxone, with clinical improvement. However, on hospitalization day 18, a sputum culture yielded carbapenem-resistant K. pneumoniae (isolate no. LC1491). She received ceftazidime/avibactam for 9 days and recovered.
Case 2
The second case occurred in a man in his mid-90s, who was admitted to the same hospital as in case 1 for weakness and turbid urine in late April 2021. His medical history included ischemic stroke with bedbound status and chronic urinary retention managed with a long-term indwelling catheter. Three months before admission, urosepsis caused by multidrug-resistant E. coli was diagnosed and treated sequentially with cefepime, meropenem, ceftazidime, and levofloxacin. He was discharged 3 weeks before the late April 2021 admission. Culture of urine collected at admission revealed carbapenem-resistant K. pneumoniae (isolate no. LC1489). He was started on ceftazidime/avibactam for urinary tract infection, but susceptibility testing revealed reduced ceftazidime/avibactam susceptibility (MIC 8 µg/mL) (Table). Treatment was then switched to fosfomycin monotherapy for 7 days, and he recovered and was discharged.
Methods
We performed antimicrobial susceptibility testing of bacterial isolates by broth microdilution, according to Clinical and Laboratory Standards Institute guidelines (16). We assessed cefiderocol susceptibility by using iron-depleted Mueller-Hinton media. We performed whole-genome sequencing on a NovaSeq platform (Illumina, https://www.illumina.com) and performed genome assembly, quality control, and identification of multilocus sequence type (ST), capsule (K locus [KL]) and O-antigen locus, resistance genes, and porin mutations, as previously described (17,18). To assess clonal relatedness among isolates, we used Snippy version 4.6 (https://github.com/tseemann/snippy) to perform core single-nucleotide polymorphism (SNP) analysis, filtering repetitive and recombination regions, as previously described (17). As the reference for core SNP analysis, we used the genome of a completely closed ST11 K. pneumoniae strain, 2020N17-130, from Taiwan (GenBank accession no. CP129835). That strain is genetically close to isolates from the 2 cases (LC1489–91) but has a wild-type PBP3.
To evaluate the functional contribution of resistance determinants, we cured the blaKPC-2 plasmid by using our previously established pCasCure plasmid curing system (19), then performed susceptibility testing of cured derivatives. To identify resistance evolution, we conducted multistep in vitro selection experiments under aztreonam/avibactam treatment by using our previously published method (20) and performed whole-genome sequencing of resistant mutants to define the underlying genetic changes. We used confocal microscopy of live/dead staining to characterize morphologic changes in K. pneumoniae carrying wild-type PBP3 and PBP3 insertions (21). In addition, we cloned blaNDM-1 into a pUC vector and introduced clones into blaKPC-2–cured strains to assess its effect on antimicrobial susceptibility profiles.
The Institutional Review Board of Taipei Veterans General Hospital provided ethics approval for the clinical data collection of both patients (approval no. 2024-01-004BC); the requirement for informed consent was waived. We deposited raw sequence data into National Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov/bioproject; BioProject no. PRJNA1308160).
Susceptibility testing showed that the 3 patient strains (LC1489–91) were resistant to imipenem and meropenem and displayed reduced susceptibility to ceftazidime/avibactam (MIC 4–8 µg/mL) and aztreonam/avibactam (MIC 1–8 µg/mL) but remained susceptible to imipenem/relebactam, meropenem/vaborbactam, and cefiderocol (Table). We next conducted conventional PCR and Sanger sequencing to examine whether the reduced ceftazidime/avibactam and aztreonam/avibactam susceptibility was caused by KPC variants (e.g., D179Y) or MBLs. Sequencing results revealed that blaKPC-2 was the only carbapenemase gene in each of the 3 isolates. The absence of KPC variants or MBLs suggested that other mechanisms caused the reduced susceptibility to ceftazidime/avibactam and aztreonam/avibactam.
Genomic analysis revealed that all 3 isolates belonged to the high-risk ST11 clone and harbored KL47 and O-antigen locus type 13. Core SNP analysis showed that the isolates differed by an average of 10 (range 8–12) SNPs, indicating a high degree of clonality. All 3 isolates carried the β-lactamase genes blaKPC-2, blaTEM-1, and blaSHV-11. The outer membrane protein OmpK35 contained a premature stop codon at amino acid position 63, and the OmpK36 protein had a glycine-aspartate insertion at position 134. Isolates LC1489 and LC1490 carried an additional plasmidborne AmpC gene, blaDHA-1 (Table). We suspect that the OmpK defects, combined with different β-lactamases, could partially contribute to the reduced aztreonam/avibactam and ceftazidime/avibactam susceptibility observed in the isolates, which is consistent with findings reported in previous studies (20,22).
Figure 1

Figure 1. Penicillin-binding protein 3 (PBP3) amino acid alignment of Klebsiella pneumoniae carbapenemase–producing K. pneumoniae with PBP3 insertions, Taiwan, 2021. Representative Escherichia coligenomes with wild-type PBP3...
Figure 2
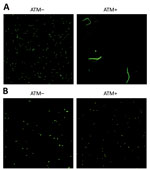
Figure 2. Confocal laser scanning microscopy of Klebsiella pneumoniae carbapenemase (KPC)–producing K. pneumoniaewith penicillin-binding protein 3 (PBP3) insertions with and without ATM treatment, Taiwan, 2021. Isolates on the...
Of note, further mining revealed that all 3 strains contained a 4–amino acid insertion, YRIT, after residue 333 (positions 334–7) in FtsI (PBP3) (Figure 1). Amino acid insertions in PBP3, particularly the tetrapeptides YRIN or YRIK at the same position (aa 334–7), frequently have been reported in E. coli and are associated with reduced susceptibility to aztreonam, cefepime, ceftazidime, and cefiderocol, all of which primarily target PBP3. However, a comparable PBP3 YRIX tetrapeptide insertion has not previously been reported in K. pneumoniae. We hypothesized that this PBP3 insertion contributed to the reduced aztreonam/avibactam and ceftazidime/avibactam susceptibility observed in those strains. Confocal laser scanning microscopy analysis showed that when treated with aztreonam (≈1/2 MICs for 6 hours), the wild-type PBP3 became elongated and filamentous (Figure 2, panel A), a typical feature of PBP3 inhibition, whereas the PBP3 with insertion mutations remained unchanged (Figure 2, panel B), suggesting that those mutations confer resistance by preventing β-lactam from effectively binding to PBP3.
Of note, on the basis of available data from E. coli (23), PBP3 tetrapeptide insertions alone are not usually sufficient to confer β-lactam resistance. Those insertions often co-occur with β-lactamases like NDM or CMY, leading to elevated resistance to aztreonam/avibactam and other β-lactams, whereas we detected KPC-2 in our 3 strains.
To assess the effects of KPC on aztreonam/avibactam susceptibility, we precisely removed the KPC plasmid, then performed susceptibility testing. Curing blaKPC-2 markedly reduced the imipenem and meropenem MICs, rendering the strains susceptible to carbapenems. However, the blaKPC-2–cured LC1489 and LC1490 strains remained resistant to aztreonam and ceftazidime because of blaDHA-1. By contrast, the blaKPC-2–cured LC1491 strain, which lacked blaDHA-1, showed an >32-fold reduction in MICs for imipenem, meropenem, aztreonam, and ceftazidime. Curing blaKPC-2 also resulted in a 2- to 4-fold decrease in the MICs of novel β-lactam/β-lactamase inhibitor combinations, including aztreonam/avibactam, ceftazidime/avibactam, meropenem/vaborbactam, and cefiderocol.
Mutations in the blaKPC gene, particularly those located in the Ω-loop, are known to cause resistance to ceftazidime/avibactam, arising through both in vitro selection and in vivo evolution. To determine whether high-level aztreonam/avibactam resistance could be obtained through blaKPC mutation, we conducted an in vitro selection experiment to evaluate whether the PBP3 YRIT insertion strain could develop high-level resistance, following a previously published protocol (20). We chose strain LC1491 for that experiment because it carried only the blaKPC-2 carbapenemase gene and the chromosome-bearing blaSHV-11. After multistep selection for aztreonam/avibactam resistance, the LC1491 strain had a MIC >128 µg/mL (Table).
To elucidate the underlying resistance mechanism, we isolated 3 resistant colonies from LC1491 (LC2124–26) and performed next-generation sequencing by using the NovaSeq platform (Illumina). Core-genome analysis revealed that the 3 mutants were nearly identical to the parental LC1491 strain, differing by <2 SNPs. Of note, all 3 resistant derivatives harbored a blaKPC-21 variant, which contained a single-nucleotide substitution (T→A at position 310) resulting in an amino acid change from tryptophan to arginine at Ambler position 105 (Trp105Arg, W105R). That variant corresponds to KPC-21, which was previously identified in a clinical E. coli ST131 isolate (24) and, in a recent in vitro selection study, was shown to confer resistance to aztreonam/avibactam in E. coli when combined with a PBP3 insertion (25).
To assess the effects of blaNDM-1 on the antimicrobial susceptibility profile of PBP3 insertions K. pneumoniae, we cloned blaNDM-1 into a pUC vector (pUC-blaNDM-1) and introduced it into blaKPC-cured LC1489, LC1490, and LC1491. The results showed that acquisition of blaNDM-1 led to high-level resistance to nearly all tested β-lactams and β-lactam/β-lactamase inhibitor combinations. Aztreonam/avibactam remained the most active agent against the 3 strains, with MICs of 4 µg/mL in pUC-blaNDM-1–harboring LC1489 and LC1490 (both co-harboring blaDHA-1) and of 1 µg/mL in pUC-blaNDM-1–harboring LC1491. In addition, cefiderocol activity was substantially reduced in the blaNDM-1 constructs, with MICs increasing to 32 µg/mL.
Taken together, findings from this study identified the emergence of K. pneumoniae ST11 strains carrying a novel 4–amino acid YRIT insertion in PBP3, which was associated with reduced susceptibility to both aztreonam/avibactam and ceftazidime/avibactam. Although PBP3 insertions have been more commonly described in E. coli, detection in K. pneumoniae suggests the possibility for dissemination of this resistance mechanism across species. Of note, K. pneumoniae ST11 is a high-risk clone capable of acquiring diverse resistance and virulence plasmids and spreading efficiently (26–28). Our findings uncover a previously unrecognized mechanism contributing to β-lactam resistance in K. pneumoniae and underscore the urgent need for continued genomic surveillance, development of novel therapeutics, and reinforced infection control measures to identify, treat, and prevent such infections.
Dr. Long is a postdoctoral researcher in the Chen Lab at the School of Pharmacy and Pharmaceutical Sciences, University at Buffalo, Buffalo, New York. His research interests focus on the molecular mechanisms of antimicrobial resistance and the development of novel therapeutics.
Acknowledgment
Research reported in this publication was supported in part by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health (award nos. R21AI190454, R01AI182297, and R01AI173064). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
References
- Sati H, Carrara E, Savoldi A, Hansen P, Garlasco J, Campagnaro E, et al.; WHO Bacterial Priority Pathogens List Advisory Group. The WHO bacterial priority pathogens list 2024: a prioritisation study to guide research, development, and public health strategies against antimicrobial resistance. Lancet Infect Dis. 2025;25:1033–43. DOIPubMedGoogle Scholar
- Rankin DA, Stahl A, Sabour S, Khan MA, Armstrong T, Huang JY, et al. Changes in carbapenemase-producing carbapenem-resistant Enterobacterales, 2019 to 2023. Ann Intern Med. 2025;178:1818–21. DOIPubMedGoogle Scholar
- Tamma PD, Heil EL, Justo JA, Mathers AJ, Satlin MJ, Bonomo RA. Infectious Diseases Society of America 2024 guidance on the treatment of antimicrobial-resistant gram-negative infections. Clin Infect Dis. 2024;8:
ciae403 . DOIPubMedGoogle Scholar - Long H, Zhao F, Feng Y, Zong Z. Global emergence of Escherichia coli with PBP3 insertions. J Antimicrob Chemother. 2025;80:178–81. DOIPubMedGoogle Scholar
- Le Terrier C, Nordmann P, Buchs C, Poirel L. Effect of modification of penicillin-binding protein 3 on susceptibility to ceftazidime-avibactam, imipenem-relebactam, meropenem-vaborbactam, aztreonam-avibactam, cefepime-taniborbactam, and cefiderocol of Escherichia coli strains producing broad-spectrum β-lactamases. Antimicrob Agents Chemother. 2024;68:
e0154823 . DOIPubMedGoogle Scholar - Sato T, Ito A, Ishioka Y, Matsumoto S, Rokushima M, Kazmierczak KM, et al. Escherichia coli strains possessing a four amino acid YRIN insertion in PBP3 identified as part of the SIDERO-WT-2014 surveillance study. JAC Antimicrob Resist. 2020;2:
dlaa081 . DOIPubMedGoogle Scholar - Alm RA, Johnstone MR, Lahiri SD. Characterization of Escherichia coli NDM isolates with decreased susceptibility to aztreonam/avibactam: role of a novel insertion in PBP3. J Antimicrob Chemother. 2015;70:1420–8. DOIPubMedGoogle Scholar
- Wang Q, Jin L, Sun S, Yin Y, Wang R, Chen F, et al. Occurrence of high levels of cefiderocol resistance in carbapenem-resistant Escherichia coli before its approval in China: a report from China CRE-Network. Microbiol Spectr. 2022;10:
e0267021 . DOIPubMedGoogle Scholar - Helsens N, Sadek M, Le Terrier C, Poirel L, Nordmann P. Reduced susceptibility to aztreonam-avibactam conferred by acquired AmpC-type β-lactamases in PBP3-modified Escherichia coli. Eur J Clin Microbiol Infect Dis. 2024;2:6. DOIPubMedGoogle Scholar
- Fabrizio C, Valzano F, Giuliano S, Morelli E, Serio D, Buccoliero GB, et al. Optimizing target inactivation to treat multidrug-resistant Escherichia coli with NDM and PBP3 mutations: “going the extra mile.”. Antimicrob Agents Chemother. 2026;20:
e0088725 . DOIPubMedGoogle Scholar - Senchyna F, Murugesan K, Rotunno W, Nadimpalli SS, Deresinski S, Banaei N. Sequential treatment failure with aztreonam-ceftazidime-avibactam followed by cefiderocol due to preexisting and acquired mechanisms in a New Delhi metallo-β-lactamase–producing Escherichia coli causing fatal bloodstream infection. Clin Infect Dis. 2024;78:1425–8. DOIPubMedGoogle Scholar
- Simner PJ, Bergman Y, Conzemius R, Jacobs E, Tekle T, Beisken S, et al. An NDM-producing Escherichia coli clinical isolate exhibiting resistance to cefiderocol and the combination of ceftazidime-avibactam and aztreonam: another step toward pan–β-lactam resistance. Open Forum Infect Dis. 2023;10:
ofad276 . DOIPubMedGoogle Scholar - Guo Y, Liu N, Lin Z, Ba X, Zhuo C, Li F, et al. Mutations in porin LamB contribute to ceftazidime-avibactam resistance in KPC-producing Klebsiella pneumoniae. Emerg Microbes Infect. 2021;10:2042–51. DOIPubMedGoogle Scholar
- Mushtaq S, Vickers A, Doumith M, Garello P, Woodford N, Livermore DM. Frequencies and mechanisms of mutational resistance to ceftibuten/avibactam in Enterobacterales. J Antimicrob Chemother. 2025;80:645–56. DOIPubMedGoogle Scholar
- Pasteran F, Manuel De Mendieta J, Pujato N, Dotta G, González LJ, Rizzo M, et al. From genomics to treatment: overcoming pan–drug-resistant Klebsiella pneumoniae in clinical settings. Front Pharmacol. 2025;16:
1570278 . DOIPubMedGoogle Scholar - Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing; thirty-fifth informational supplement (M100-S35). Wayne (PA): The Institute; 2025.
- van Duin D, Arias CA, Komarow L, Chen L, Hanson BM, Weston G, et al.; Multi-Drug Resistant Organism Network Investigators. Molecular and clinical epidemiology of carbapenem-resistant Enterobacterales in the USA (CRACKLE-2): a prospective cohort study. Lancet Infect Dis. 2020;20:731–41. DOIPubMedGoogle Scholar
- Wang M, Earley M, Chen L, Hanson BM, Yu Y, Liu Z, et al.; Multi-Drug Resistant Organism Network Investigators. Clinical outcomes and bacterial characteristics of carbapenem-resistant Klebsiella pneumoniae complex among patients from different global regions (CRACKLE-2): a prospective, multicentre, cohort study. Lancet Infect Dis. 2022;22:401–12. DOIPubMedGoogle Scholar
- Hao M, He Y, Zhang H, Liao XP, Liu YH, Sun J, et al. CRISPR-Cas9–mediated carbapenemase gene and plasmid curing in carbapenem-resistant Enterobacteriaceae. Antimicrob Agents Chemother. 2020;64:e00843–20. DOIPubMedGoogle Scholar
- Niu S, Wei J, Zou C, Chavda KD, Lv J, Zhang H, et al. In vitro selection of aztreonam/avibactam resistance in dual-carbapenemase-producing Klebsiella pneumoniae. J Antimicrob Chemother. 2020;75:559–65. DOIPubMedGoogle Scholar
- Lang Y, Shah NR, Tao X, Reeve SM, Zhou J, Moya B, et al. Combating multidrug-resistant bacteria by integrating a novel target site penetration and receptor binding assay platform into translational modeling. Clin Pharmacol Ther. 2021;109:1000–20. DOIPubMedGoogle Scholar
- Mendes RE, Doyle TB, Streit JM, Arhin FF, Sader HS, Castanheira M. Investigation of mechanisms responsible for decreased susceptibility of aztreonam/avibactam activity in clinical isolates of Enterobacterales collected in Europe, Asia and Latin America in 2019. J Antimicrob Chemother. 2021;76:2833–8. DOIPubMedGoogle Scholar
- Ma K, Feng Y, McNally A, Zong Z. Struggle to survive: the choir of target alteration, hydrolyzing enzyme, and plasmid expression as a novel aztreonam-avibactam resistance mechanism. mSystems. 2020;5:e00821–20. DOIPubMedGoogle Scholar
- Manageiro V, Romão R, Moura IB, Sampaio DA, Vieira L, Ferreira E, et al. Network EuSCAPE-Portugal. Molecular epidemiology and risk factors of carbapenemase-producing Enterobacteriaceae isolates in Portuguese hospitals: results from European Survey on Carbapenemase-Producing Enterobacteriaceae (EuSCAPE). Front Microbiol. 2018;9:2834. DOIPubMedGoogle Scholar
- Ma K, Feng Y, Zong Z. Aztreonam-avibactam may not replace ceftazidime/avibactam: the case of KPC-21 carbapenemase and penicillin-binding protein 3 with four extra amino acids. Int J Antimicrob Agents. 2022;60:
106642 . DOIPubMedGoogle Scholar - Dong N, Yang X, Chan EW, Zhang R, Chen S. Klebsiella species: taxonomy, hypervirulence and multidrug resistance. EBioMedicine. 2022;79:
103998 . DOIPubMedGoogle Scholar - Yang X, Dong N, Chan EW, Zhang R, Chen S. Carbapenem resistance–encoding and virulence-encoding conjugative plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021;29:65–83. DOIPubMedGoogle Scholar
- Yang Y, Qin J, Hu Y, Wang J, Feng Y, Zong Z. Carbapenem-resistant Klebsiella pneumoniae of sequence type 11: a scoping review. J Infect Dis. 2026;233(Supplement_1):S72–80. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 32, Number 6—June 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Liang Chen, School of Pharmacy and Pharmaceutical Sciences, University at Buffalo, 360 Hayes Rd, Buffalo, NY 14214, USA
Top