Volume 14, Number 12—December 2008
Dispatch
Multiple Francisella tularensis Subspecies and Clades, Tularemia Outbreak, Utah
Cite This Article
Citation for Media
Abstract
In July 2007, a deer fly–associated outbreak of tularemia occurred in Utah. Human infections were caused by 2 clades (A1 and A2) of Francisella tularensis subsp. tularensis. Lagomorph carcasses from the area yielded evidence of infection with A1 and A2, as well as F. tularensis subsp. holarctica. These findings indicate that multiple subspecies and clades can cause disease in a localized outbreak of tularemia.
Tularemia is a zoonotic disease caused by Francisella tularensis, a highly infectious, gram-negative coccobacillus found in lagomorphs (rabbits and hares), rodents, and arthropods throughout the Northern Hemisphere. Humans become infected through contact with infected animal tissues, ingestion of contaminated food or water, inhalation of contaminated aerosols, and bites of arthropods, especially ticks and deer flies.
In North America, tularemia is caused by 2 subspecies of F. tularensis, subsp. tularensis (type A) and subsp. holarctica (type B). The distribution of type A and type B strains appears largely overlapping within the United States, with some geographic distinctions (1,2). Ecologically, the 2 subspecies are thought to be maintained in distinct but incompletely defined cycles, with type A strains frequently associated with lagomorphs and type B strains more commonly associated with rodents and aquatic environments (3).
Type A strains can be further divided into 2 major clades by various molecular subtyping techniques (1,2,4–7). These clades, designated here as A1 and A2, differ in their overall geographic distribution and clinical features. A1 strains (also known as A.I. and A-east) are usually found east of the Rocky Mountains. A2 strains (also known as A.II. and A-west) are common in the intermountain region of the western United States, are associated with lower mortality rates in humans, and are the only strains currently linked to transmission by deer flies (Chrysops spp.) (2).
Figure 1

Figure 1. Dendrogram based on PmeI pulsed-field gel electrophoresis (PFGE) patterns of Francisella tularensis type A isolates. The dendrogram was constructed by using Dice similarity coefficients (1.5% optimization and 1.5% tolerance) and unweighted...
In July 2007, an outbreak of ulceroglandular tularemia occurred in Utah among visitors to the southwest shore of Utah Lake; an epidemiologic investigation implicated deer fly bites as the source of infection (8). Clinical isolates were obtained July 9–14 from skin lesions of 5 patients. Isolates were identified as F. tularensis subsp. tularensis (type A) by biochemical analysis (glycerol fermentation). Molecular subtyping of isolates was performed by using PmeI pulsed-field gel electrophoresis (PFGE) as previously described (2). PFGE gels were normalized by comparison to the Salmonella enterica serotype Braenderup (H9812) reference strain by using BioNumerics software (v. 4.0, Applied Maths BVBA, Sint-Martens-Latem, Belgium) (2). A dendrogram was constructed by comparison with PmeI PFGE patterns for A1 (SCHU S4, MA00–2972) and A2 (ATCC 6223, WY96–3418) control strains (1,2,4–7,9) (Figure 1). The 5 clinical isolates fell into the 2 major type A clades; 2 isolates were identified as A2 (UT07–4632, UT07–4633), and 3 were identified as A1 (UT07–4262, UT07–4263, UT07–4265) (Figure 1). PFGE patterns for the 3 A1 isolates were indistinguishable from each other, as were patterns for the 2 A2 isolates (Figure 1).
Figure 2
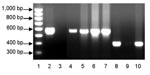
Figure 2. PCR typing of Francisella tularensis, clades A1 and A2, in dessicated lagomorph carcasses. Lane 1, 100-bp ladder (Bio-Rad, Hercules, CA, USA); lane 2, A1 positive control (Schu S4); lane 3, A1...
A brief search of the exposure area yielded desiccated carcasses of 10 black-tailed jackrabbits (Lepus californicus) and 2 desert cottontail rabbits (Sylvilagus audubonii). The carcasses were found within a few hundred meters of each other, in an overall area <0.8 km across. Living deer flies (Chrysops spp.) were collected from the same area. DNA was extracted from lagomorph bone marrow and flies by using the QIAamp DNA MiniKit (QIAGEN, Valencia, CA, USA) and tested with real-time PCR F. tularensis multitarget, type A and type B assays (10,11). Although all deer fly samples were negative, 11 of 12 lagomorph carcasses tested positive for F. tularensis by the multitarget assay (3 of 3 targets positive; crossing threshold (Ct) range 13–38). Among the infected samples, 9 tested positive for type A (Ct range 13–36) and 2 tested positive for type B (Ct range 19–24). The subtyping results were verified by sequencing of the succinate dehydrogenase gene (sdhA), which distinguishes type A and type B strains on the basis of a single nucleotide polymorphism (12). Type B strains have a G at nt 465 of the sdhA gene sequence, whereas type A strains have an A at this position. To further distinguish the type A samples between clade A1 or clade A2, conventional PCRs were used (13). Sufficient F. tularensis DNA was present to type infections for 5 of the 9 type A–positive lagomorph carcasses; 4 yielded a PCR product consistent with the A1 clade (570 bp), and 1 yielded a product consistent with the A2 clade (396 bp) (Figure 2) (13).
Few studies have reported on the diversity of F. tularensis subsp. or clades present during outbreaks of tularemia, in part because molecular methods for strain discrimination have only recently been described (1,2,4–7). In this discrete deer fly–associated outbreak, we found human infections caused by both A1 and A2 strains, and evidence that A1, A2, and type B strains were circulating among lagomorphs in the exposure area. These findings demonstrate that multiple subspecies and clades can cause disease in a localized outbreak of tularemia and that deer-flies are associated with transmission of A1 strains.
Published reports indicate that A1 and A2 strains are generally segregated into areas east and west of the Rocky Mountains, respectively, with some overlap in coastal California (1,2). In contrast, our results demonstrate that A1 strains are present in areas of the intermountain west and at elevations >1,200 m. This finding is supported by identification of an additional case of human tularemia in Utah caused by an A1 strain in 1998 (Centers for Disease Control and Prevention, unpub. data). On a local level, our results indicate that A1, A2, and type B strains can coexist naturally within the same ecosystem, a paradox when compared with the segregation that appears to exist on a larger scale. Overall, these observations underscore the need for future studies to define the ecologic and evolutionary factors underlying the distributions of F. tularensis strains in North America.
Although the role of deer flies as vectors of F. tularensis is well established, the dynamics of deer fly–associated outbreaks have not been well researched. Transmission of F. tularensis by deer flies is believed to be entirely mechanical, through contamination of the mouthparts. Long-term maintenance of F. tularensis has not been shown to occur in deer flies, and it is therefore not surprising that the deer flies we collected 3 weeks after the outbreak tested negative for this organism. Our findings suggest that deer flies nonselectively acquire and transmit whatever strains are circulating in enzootic hosts. We postulate that, in this instance, an abundance of deer flies led to extensive feeding on many hosts, resulting in the simultaneous transmission of multiple strains. High mortality rates among lagomorphs may have forced deer flies to seek alternate hosts, specifically muskrats, which are associated with type B strains and have been linked to outbreaks among trappers at Utah Lake (3).
The co-occurrence of multiple subspecies and clades may be unique to arthropod-associated outbreaks of tularemia and not characteristic of outbreaks resulting from other modes of F. tularensis transmission, such as contaminated water. Further work is needed to determine whether our findings will apply to other deer fly–associated outbreaks or for outbreaks of tularemia associated with ticks, which are known to maintain as well as transmit F. tularensis. Notably, while investigating a tick-borne outbreak of presumed type B infections in South Dakota, Markowitz and colleagues found evidence of both type A and type B strains in Dermacentor variabilis ticks collected from dogs (14). Outbreaks involving multiple serotypes have been observed with other vector-borne pathogens, including dengue virus (15), which suggests that amplification and transmission of multiple strains in a focal area may represent a general feature of some vector-borne diseases.
Dr Petersen is a research microbiologist with the Bacterial Zoonoses Branch, Division of Vector-Borne Infectious Diseases, Centers for Disease Control and Prevention, Fort Collins, Colorado. Her research interests focus on improving diagnosis of tularemia to gain a better understanding of the ecology and epidemiology of the disease in North America.
Acknowledgment
We thank Robert Rolfs, Kim Christensen, John Amadio, James Colburn, Ken Gage, Kiersten Kugeler, Marty Schriefer, Sandy Urich, and John Young, and the staff of the Salt Lake Valley and Utah County Departments of Health for their assistance.
References
- Farlow J, Wagner DM, Dukerich M, Stanley M, Chu M, Kubota K, Francisella tularensis in the United States. Emerg Infect Dis. 2005;11:1835–41.PubMedGoogle Scholar
- Staples JE, Kubota KA, Chalcraft LG, Mead PS, Petersen JM. Epidemiologic and molecular analysis of human tularemia, United States, 1964–2004. Emerg Infect Dis. 2006;12:1113–8.PubMedGoogle Scholar
- Jellison WL. Tularemia in North America, 1930–1974. Missoula (MT): University of Montana; 1974.
- Johansson A, Farlow J, Larsson P, Dukerich M, Chambers E, Bystrom M, Worldwide genetic relationships among Francisella tularensis isolates determined by multiple-locus variable-number tandem repeat analysis. J Bacteriol. 2004;186:5808–18. DOIPubMedGoogle Scholar
- Svensson K, Larsson P, Johansson D, Byström M, Forsman M, Johansson A. Evolution of subspecies of Francisella tularensis. J Bacteriol. 2005;187:3903–8. DOIPubMedGoogle Scholar
- Larsson P, Svensson K, Karlsson L, Guala D, Granberg M, Forsman M, Canonical insertion-deletion markers for rapid DNA typing of Francisella tularensis. Emerg Infect Dis. 2007;13:1725–32.PubMedGoogle Scholar
- Nübel U, Reissbrodt R, Weller A, Grunow R, Porsch-Ozcürümez M, Tomaso H, Population structure of Francisella tularensis. J Bacteriol. 2006;188:5319–24. DOIPubMedGoogle Scholar
- Tularemia–USA (Utah. New Jersey) (02). Promed. 2007 Jul 18 [cited 2008 Oct 1]. Available from http://www.promedmail.org, archive no. 20070718.2304.
- Beckstrom-Sternberg SM, Auerbach RK, Godbole S, Pearson JV, Beckstrom-Sternberg JS, Deng Z, Complete genomic characterization of a pathogenic A.II strain of Francisella tularensis subspecies tularensis. PLoS One. 2007;2:e947. DOIPubMedGoogle Scholar
- Versage JL, Severin DD, Chu MC, Petersen JM. Development of a multitarget real-time TaqMan PCR assay for enhanced detection of Francisella tularensis in complex specimens. J Clin Microbiol. 2003;41:5492–9. DOIPubMedGoogle Scholar
- Kugeler KJ, Pappert R, Zhou Y, Petersen JM. Real-time PCR for Francisella tularensis types A and B. Emerg Infect Dis. 2006;12:1799–801.PubMedGoogle Scholar
- Barns SM, Grow CC, Okinaka RT, Keim P, Kuske CR. Detection of diverse new Francisella-like bacteria in environmental samples. Appl Environ Microbiol. 2005;71:5494–500. DOIPubMedGoogle Scholar
- Molins-Schneekloth CR, Belisle JT, Petersen JM. Genomic markers for differentiation of Francisella tularensis subsp. tularensis A.I and A.II strains. Appl Environ Microbiol. 2008;74:336–41. DOIPubMedGoogle Scholar
- Markowitz LE, Hynes NA, de la Cruz P, Campos E, Barbaree JM, Plikaytis BD, Tick-borne tularemia. An outbreak of lymphadenopathy in children. JAMA. 1985;254:2922–5. DOIPubMedGoogle Scholar
- Fang R, Lo E, Lim TW. The 1982 dengue epidemic in Malaysia: epidemiological, serological and virological aspects. Southeast Asian J Trop Med Public Health. 1984;15:51–8.PubMedGoogle Scholar
Figures
Cite This ArticleTable of Contents – Volume 14, Number 12—December 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Paul S. Mead, Centers for Disease Control and Prevention, Division of Vector-Borne Infectious Diseases, 3150 Rampart Rd, Fort Collins, CO 80521, USA
Top