Volume 14, Number 12—December 2008
CME ACTIVITY - Research
Clinical Characteristics and Molecular Subtyping of Vibrio vulnificus Illnesses, Israel
Cite This Article
Citation for Media
Introduction
![]()
Medscape, LLC is pleased to provide online continuing medical education (CME) for this journal article, allowing clinicians the opportunity to earn CME credit. Medscape, LLC is accredited by the Accreditation Council for Continuing Medical Education (ACCME) to provide CME for physicians. Medscape, LLC designates this educational activity for a maximum of 0.75 AMA PRA Category 1 Credits™. Physicians should only claim credit commensurate with the extent of their participation in the activity. All other clinicians completing this activity will be issued a certificate of participation. To participate in this journal Medscape CME activity: (1) review the learning objectives and author disclosures; (2) study the education content; (3) take the post-test and/or complete the evaluation at http://www.medscape.com/cme/eid; (4) view/print certificate.
Learning Objectives
- Upon completion of this activity, participants will be able to:
- List the predisposing factors for infections caused by Vibrio vulnificus biotypes 1 and 2.
- Identify the differences between infections caused by V. vulnificus biotypes 1 and 2 and biotype 3.
- Describe the types of fish associated with V. vulnificus biotype 3 infection.
- Describe the mortality associated with V. vulnificus biotype 3 infection.
- List the predictors of mortality in V. vulnificus biotype 3 infection.
Medscape CME Editor
Beverly Merritt, Technical Writer-Editor, Emerging Infectious Diseases. Disclosure: Beverly Merritt has disclosed no relevant financial relationships.
Medscape CME Author
Désirée Lie, MD, MSEd, Clinical Professor, Family Medicine, University of California, Orange; Director, Division of Faculty Development, UCI Medical Center, Orange, California. Disclosure: Désirée Lie, MD, MSEd, has disclosed no relevant financial relationships.
Authors
Disclosures: Ronit Zaidenstein, MD; Chantal Sadik, MD; Larisa Lerner, MD; Lea Valinsky, PhD; June Kopelowitz, PhD; Ruth Yishai, PhD; Vered Agmon, PhD; Michele Parsons, MS; Cheryl Bopp, MS; and Miriam Weinberger, MD, have disclosed no relevant financial relationships.
Abstract
During 1996–1997, a new Vibrio vulnificus biotype 3, which caused severe soft tissue infection after fishbone injury, emerged in Israel. We conducted a follow-up study from 1998 through 2005 to assess changing trends, outcomes, and molecular relatedness of the implicated strains. A total of 132 cases (71% confirmed and 29% suspected) of V. vulnificus biotype 3 infection were found. Most infections (95%) were related to percutaneous fish exposure, mainly tilapia (83%) or common carp (13%). Bacteremia, altered immune status, and history of ischemic heart disease were identified as independent risk factors for death, which reached a prevalence of 7.6%. Pulsed-field gel electrophoresis patterns of strains from 1998 through 2005 and from 1996 through 1997 showed a high degree of homogeneity and were distinct from those of V. vulnificus biotype 1. Infections caused by V. vulnificus biotype 3 continue affect the public’s health in Israel.
Vibrio vulnificus, a gram-negative bacterium of the family Vibrionaceae, is a worldwide inhabitant of salt water (1–3). V. vulnificus biotypes 1 and 2 are capable of causing severe human infection, including necrotizing fasciitis and septicemia; the death rate is substantial (4–6). Persons with chronic liver disease, particularly liver cirrhosis, are more prone to developing infection and at greatest risk for an adverse outcome (7,8). Other predisposing factors are iron overload and hemochromatosis and immunosuppression caused by steroid treatment, malignancy, HIV infection, renal failure, and organ transplantation (1,9,10).
During 1996–1997, a new biotype, V. vulnificus biotype 3, emerged as a cause of severe soft tissue infection and bacteremia in Israel (11,12). Several important features differentiate the illness caused by the new V. vulnificus biotype from previously described V. vulnificus infections. First, a new vector, a pond fish (tilapia) grown in fresh water, has been associated with V. vulnificus infection. Second, infection is caused by direct injury from the fish backbone while purchasing, cleaning, or handling the live fish, as opposed to contamination of a prior injury by immersion in seawater or ingestion of contaminated seafood. Bacteriologically, the new V. vulnificus biotype differs from other V. vulnificus strains by its biochemical features (salicin-, cellobiose-, citrate-, and lactose-negative, plus delayed reaction for o-nitrophenyl-β-
The 1996–1997 Israeli cluster involved 62 persons, with a slight male predominance (58%) and a median age of 56 years. Although no deaths were reported, 41 persons (66%) had conditions that required surgical debridement, 1 had total limb amputation, and 7 had finger amputations (11). A new, aggressive, live-fish marketing initiative in the northern part of Israel was implicated in the outbreak, and the outbreak was followed by new instructions from the Israeli Ministry of Health that recommended selling only precleaned, ice-chilled tilapia (17).
We studied the epidemiology and the trends in illnesses associated with the new V. vulnificus biotype 3 during a 7-year period (1998–2005) following the initial 1996–1997 cluster. Our study assesses the effects of infection, risk factors for death, possible spread to other fish species, and molecular relatedness of the V. vulnificus biotype 3 strain.
Clinical Data
Clinical data were obtained from the records of the Infectious Diseases Department, Epidemiology Unit, Israel Ministry of Health, Jerusalem. Subdistrict health offices reported data collected from persons with V. vulnificus infections; a standardized questionnaire was used. Investigation was initiated after passive reporting from primary physicians who treated patients with suspected V. vulnificus infection or when clinical isolates were positively identified as V. vulnificus biotype 3 by the Vibrio Reference Laboratory in the Government Central Laboratories, Israel Ministry of Health, Jerusalem. The data collected were patient’s age, sex, and place of residence; underlying diseases; circumstances of exposure and type of fish involved; time lapse from exposure to seeking treatment in the emergency department; site of infection; length of hospitalization; clinical symptoms; source of isolation; antimicrobial drug treatment; and outcome.
Cases were classified as laboratory-confirmed when a patient with suggestive history had V. vulnificus or V. vulnificus biotype 3 isolated from blood or soft tissue or as suspected when a patient had suggestive history without positive cultures. Suggestive history was considered to be the development of soft tissue infection or septicemia after recent (within 7 days) fish exposure or immersion in a water pool.
Source of Isolates and Laboratory Diagnosis
Initial identification of V. vulnificus was performed in the microbiology laboratories of hospitals where infected patients had been admitted, using the API 20E strip (bioMérieux, Marcy-l’Etoile, France). The laboratories submitted the isolates for confirmation and further identification to the Vibrio Reference Laboratory, Government Central Laboratories in Jerusalem. Identification of V. vulnificus biotype 3 was performed solely in this laboratory by using biochemical tests (failure to ferment citrate, lactose, salicin, cellobiose, and a negative test result for ONPG).
Figure 1
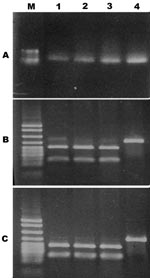
Figure 1. PCR restriction fragment length polymorphism of Vibrio vulnificus cytotoxin gene vvhA. A) PCR amplicon of vvhA gene restriction digested with B) PstI or C) KpnI. Gel shows molecular size standards (M)...
Random isolates were also submitted for molecular analysis of the cytotoxin-hemolysin gene. This test was developed in the Vibrio Reference Laboratory and can differentiate between V. vulnificus biotype 3 and biotypes 1 and 2 by demonstrating the unique restriction fragment length polymorphism (RFLP) patterns of biotype 3 (Figure 1). Briefly, the V. vulnificus cytotoxin-hemolysin gene (vvhA) is amplified from crude bacteria lysate (boiling a loop full of bacteria suspended in 100 μL Tris-EDTA buffer for 10 min) using primers with the sequences RRCTH 5′-CAGCTCCAGCCGTTAACCGAACCACCCGC-3′ and LCTH 5′-TTCCAACTTCAAACCGAACTATGAC-3′. This step is followed by RFLP analysis. For the enzymatic restriction reaction, 2 μL of the PCR-amplified DNA was added to a reaction mixture to give a final volume of 20 μL, according to the manufacturer’s instructions (New England Biolabs, Ipswich, MA, USA), in 2 separate reactions. The restricted DNA was separated by electrophoresis in a 2% gel that was stained with ethidium bromide and visualized for the specific biotype 3 restriction patterns. Restriction sites for 2 enzymes, KpnI and PstI, existed in the sequence of vvhA of V. vulnificus biotype 3 but not in the corresponding vvhA gene of biotypes 1 and 2 (Figure 1).
Molecular Subtyping
Twenty randomly selected laboratory-confirmed isolates of V. vulnificus biotype 3 from study years (1998–2003) plus 4 retrospective isolates from 1997 were sent to Centers for Disease Control and Prevention (CDC), Atlanta, Georgia, USA, for molecular subtyping. The isolates were recovered from the blood or wound sites of patients with a history of exposure to various fish. An additional isolate was recovered from the remnants of a tilapia fish found in the refrigerator of 1 of the infected patients (the fish remnants were diced and inoculated onto Vibrio-selective agar with and without enrichment). These 25 isolates were available for further molecular studies. Eight submitted human V. vulnificus biotype 1 isolates stored at CDC were used for comparison.
V. vulnificus isolates were subtyped by pulsed-field gel electrophoresis (PFGE) in accordance with the PulseNet protocol for V. cholerae (18) using SfiI for primary enzyme digestion with 1 modification: thiourea (Sigma-Aldrich, St. Louis, MO, USA) was routinely added to the electrophoresis running buffer at a final concentration of 50 μmol/L (19) to prevent DNA degradation, which was commonly found during the initial runs. All DNA fingerprints were captured using a Gel Doc EQ system (Bio-Rad, Hercules, CA, USA). The PFGE fingerprints were analyzed in BioNumerics, version 4.0 (Applied Maths, Sint-Martens-Latem, Belgium). Gels were normalized by aligning the bands of the PulseNet universal standard Salmonella enterica serotype Braenderup (H9812) placed in every fifth lane on the gels (20). Dendrograms were made of the similarities of the DNA fingerprints by using the Dice similarity coefficient and unweighted pair group method with averages (unweighted pair group method with arithmetic mean) clustering. An optimization of 1.5% and tolerance window of 1.5% were used. The 25 isolates sent to CDC as described above were submitted for susceptibility tests using the E-test method according to Clinical Laboratory Standards Institute standards for Enterobacteriaceae (21).
Statistical Methods
Proportions were compared by using the Fisher exact test or the χ2 test, and continuous variables were compared by using the Kruskal-Wallis test. Variables associated with death at the 0.05 significance level were entered into a stepwise forward logistic regression model for mortality rate. Analyses were performed by using SPSS version 15 software (SPSS, Inc., Chicago, IL, USA).
Figure 2
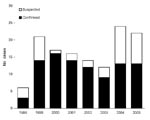
Figure 2. Annual distribution of laboratory-confirmed and suspected Vibrio vulnificus biotype 3 infections.
Figure 3
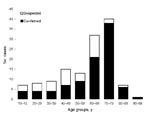
Figure 3. Age distribution of patients with laboratory-confirmed and suspected Vibrio vulnificus biotype 3 infections.
A total of 134 cases of V. vulnificus infection were identified during the 8-year study period from 1998 through 2005 (Figure 2). Most cases (96, 71.6%) were laboratory-confirmed; 70 (52%) were submitted to the Vibrio Reference Laboratory and identified as V. vulnificus biotype 3. PCR-RFLP analysis performed on 34 of these 70 isolates (49%) showed the unique KpnI, PstI pattern. Two patients with laboratory-confirmed V. vulnificus biotype 3 infection were excluded from further analyses because clinical data were missing. The median age of the remaining 132 patients was 66 years (mean 58.9 years, range 10–93 years) (Figure 3). Overall, infection rates for women were only slightly higher than those for men (1.1:1); this predominance increased in those >70 years of age (1.7:1). The sites of V. vulnificus isolation included wounds in 61 patients (65% of 94 patients with laboratory-confirmed infection), blood in 24 (26%), or both in 7 (7%). The source was not recorded in 2 patients.
Clinical characteristics of the patients and outcomes are summarized in Table 1. Compared to patients with laboratory-confirmed infection, patients with suspected infection were more likely to be men, with a longer incubation period after exposure, less severe clinical symptoms, a shorter hospitalization time, and a more favorable outcome. Information regarding the type of exposure was available for 93 patients. Most of these patients (75, 81%) were injured while purchasing or preparing fish for cooking. Fish were purchased from fish stores (52, 56%), mobile selling units (9, 10%), or stands near fishponds (14, 15%). Other types of exposures included involvement in fish marketing (14, 15%), either in selling (11/14) or cleaning the fishponds (3/14). One person was infected after fishing at a fishpond. Four persons had no connection to the pond fish industry. Three persons with suspected infection became ill after fishing in the Sea of Galilee, and 1 with laboratory-confirmed V. vulnificus biotype 3 infection reported immersion in a natural spring near Jerusalem with an open wound. Most of the injuries affected the hands; only 9 affected the legs.
Figure 4

Figure 4. Seasonality of Vibrio vulnificus biotype 3 illnesses, Israel, 1998–2005.
Most of the infections (74%) occurred in the warm months of the year between June and November (Figure 4). Most patients resided in northern Israel, where freshwater fish aquaculture takes place. Only 6 patients lived in other areas of Israel (3 in the central part, 2 in the southern part, and 1 near Jerusalem). A total of 104 patients provided details regarding fish exposure. Tilapia (St. Peter’s fish) was the primary fish involved in 86 of these cases (83%), common carp (Cyprinius carpio) in 14 cases (13%), and both in 2 cases (2%). Two other patients reported exposure to saltwater fish: gilt-head sea brim (Sparus aurata) that was purchased in a fish store (laboratory-confirmed infection) and common gray mullet (Mugil cephalous) that was purchased at a fish stand near a fishpond (suspected infection).
Course and Outcome
A total of 124 patients (94%) were hospitalized for a mean duration of 10.9 days (median 9 days, range 2–50 days). Information regarding treatment with antimicrobial drugs was available for 112 patients and included a variety of regimens. The most frequent regimens among these patients included only a single drug, usually ampicillin-clavulanate (47 patients, 42%), doxycycline (27, 24%), and ceftriaxone (6, 5%). Combinations of drugs were also used and included third-generation cephalosporins (ceftriaxone or ceftazidime) plus doxycycline (18, 16%), third-generation cephalosporins plus ampicillin-clavulanate (3, 3%), or doxycycline plus ampicillin-clavulanate (8, 7%).
Ten patients (7.5%) died and 9 (6.8%) underwent amputation of fingers or part of the arm (7 patients) or a leg (2). Two of the patients who underwent amputation subsequently died. Seven of the patients who died (70%) were >70 years of age, 2 were in their 50s, and 1 was in his 60s. All patients who died or underwent amputation had laboratory-confirmed infection.
Ninety patients with laboratory-confirmed infections were included in the analyses of risk factors associated with death. Factors that were significantly associated with death by a univariate analysis are outlined in Table 2. We could not show that any of the antimicrobial drug regimens influenced outcomes. Variables associated with death were entered into a stepwise forward logistic regression model. All initially entered variables remained in the model as independent predictors of a fatal outcome: presence of bacteremia (odds ratio [OR] 6.03, 95% confidence interval [CI] 1.2–30.8, p = 0.031), altered immune status (OR 6.7, 95% CI 1.1–40.8, p = 0.038), and history of ischemic heart disease (OR 15, 95% CI 2.5–91.1, p = 0.003) (p value for the model <0.001). When septic shock was added, only septic shock (OR 24.4, 95% CI 4.3–140.2, p<0.001) and history of ischemic heart disease (OR 9.7, 95% CI 1.3–72.7, p = 0.027) remained in the model (p value for the model <0.001). The model did not change when adjusted for age.
Antimicrobial Drug Susceptibility Tests
V. vulnificus isolates were susceptible to all antimicrobial agents tested by criteria for Enterobacteriaceae. The MIC ranges for ampicillin were 0.5–1.0 μg/mL; cephalothin, 4–8 μg/mL; chloramphenicol, 0.5–0.75 μg/mL; ciprofloxacin, 0.012–0.023 μg/mL; kanamycin, 4–16 μg/mL; nalidixic acid, 0.25–1 μg/mL; streptomycin, 8–16 μg/mL; tetracycline, 0.5–7.5 μg/mL; and trimethoprim-sulfamethoxazole, 0.064–0.094 μg/mL.
Molecular Subtyping
Figure 5

Figure 5. Dendogram comparing pulsed-field gel electrophoresis patterns of 25 Vibrio vulnificus biotype 3 isolates and a reference set of biotype 1 isolates when restricted with SfiI enzyme.
Analysis of the PFGE subtyping results from 25 isolates (21 isolates from 1998 through 2003, 4 isolates from 1997) showed that the selected isolates represented 18 unique but similar (>88% pattern similarity; 1–3 fragment differences) PFGE fingerprint patterns regardless of the type of fish exposure (tilapia vs. carp) and year of isolation (Figure 5). A tilapia isolate from 2003 generated an SfiI PFGE pattern, which was indistinguishable from the PFGE patterns of 3 patient isolates (1 from 2002, 2 from 2003) with reported exposure to tilapia from wound sites. When compared with biotype 1 strains reported from the United States, the biotypes separated into 2 distinct clusters with ≈70% pattern similarity (Figure 5).
Our study showed that infections caused by V. vulnificus biotype 3 continued to occur after the initial cluster during 1996–1997, with an average of ≈16 cases annually. Although the annual rate of infection during 1998–2005 was half the rate of infection during 1996–1997 (62 cases) (11), outcomes were more grave. Our study found that 10 persons (7.6%) died; no deaths were reported during the 1996–1997 outbreak (11). Possible explanations for this disparity could be that patients in our study were older (median age 66 vs. 56 years, respectively) with a higher proportion of laboratory-confirmed infections (70.1% vs. 53%, respectively). We have shown that patients without laboratory confirmation had a much milder form of the disease and a more favorable outcome.
Notably, our findings show a high proportion of infected women (52%), including 8 (80%) of the 10 patients who died. Previous studies have stressed a male predominance (7,9,10,22–25), and have even argued that female sex hormones protect against contracting the disease (26). With V. vulnificus biotype 3 infections in Israel, it was more likely that women purchased and prepared the fish before cooking, and thus were more likely to be exposed to fishbone injuries.
There are 3 major clinical syndromes of V. vulnificus illnesses, including primary bacteremia (mostly related to raw seafood consumption), wound infection (mostly related to immersion in contaminated water or to injury by seafood preparation), and gastroenteritis (after consumption of seafood or swallowing contaminated water) (9,23,27). Patients with primary bacteremia caused by non–biotype 3 strains are more likely to have predisposing conditions, particularly liver diseases in >80% of patients (7–9), whereas patients with wound infection are more likely to be previously healthy and have a more favorable outcome (9,10,23,24,28–30). The death rate may exceed 50% in the most seriously ill patients (7–9,23).
Our study describes a large, uniform group of patients who acquired infection through percutaneous exposure. All sought treatment for a wound infection, which in 18% was complicated by secondary bacteremia. In 38 patients (29%), laboratory confirmation of infection was lacking. These patients tended to delay seeking treatment and to have a milder form of infection, compared with patients who had laboratory-confirmed infection. Some of the patients who lacked laboratory confirmation may not have had V. vulnificus infection. However, patients with milder forms of V. vulnificus infection may have been less likely to undergo extensive microbiologic workup and the yield of cultures, if taken, would have been lower.
Only 45% of the patients in the group had underlying diseases. Liver disease occurred in 14% of patients overall, and in 19% of patients with confirmed cases, but was not a statistically significant risk factor for death. The case-fatality rate was 7.6% for the entire study population and 10.6 % for patients with laboratory-confirmed infection. These characteristics concur with prior reports of wound infection syndrome caused by non–biotype 3 V. vulnificus (9,10,23,24,27–30).
Of concern is our finding that V. vulnificus infection was not limited to tilapia exposure. Fourteen persons (13%) reported exposure to the common carp, or to both tilapia and common carp (2 persons, 2%). Tilapia and common carp are co-cultivated in several fishponds in northern Israel, which may have resulted in cross-contamination. For those exposed to salt water fish (gilt-head sea brim and common gray mullet, 1 person each), contamination may have occurred at the place of purchase. V. vulnificus infection after exposure to the common carp caused the death of 1 patient and the amputation of a finger in another. One person infected after exposure to the gilt-head sea brim also died. Notably, orthopedic surgeons from hospitals in northern Israel have also pointed out severe soft tissue infections, including those caused by V. vulnificus, after fishbone injury from the common carp (31).
The V. vulnificus biotype 3 strains studied were uniformly susceptible to all tested antimicrobial agents. These results concur with those of similar studies of clinical and environmental V. vulnificus biotype 1 isolates from the United States (32,33) and Taiwan (22). Only a few isolates from Taiwan showed resistance to ceftazidime and moxalactam.
Molecular subtyping of the V. vulnificus biotype 3 strains by PFGE showed no specific association between fish species and PFGE pattern. The results indicate that the biotype 3 strains are homogeneous with limited heterogeneity between the isolates but cluster distinct from biotype 1 strains. PFGE has been shown (34) to offer sufficient discrimination when subtyping biotype 1 strains, but published findings that evaluate the utility of PFGE to sufficiently discriminate between biotype 3 strains are limited. Modifications in restriction sites may alter the number and size of DNA fragments, which define the PFGE pattern and result in observable true differences.
A high degree of homogeneity among the V. vulnificus biotype 3 strains and distinction from the biotypes 1 and 2 has been also observed by other authors applying various methods, including random amplified polymorphic DNA, (13), multilocus sequence typing (14,16), and analysis of variations in simple sequence repeat loci (35). Notably, the latter method was able to demonstrate small-scale variations among the biotype 3 strains (35). The PFGE results also support the conclusion that this biotype 3 is distinct from the other V. vulnificus biotypes. The high degree of homogeneity is another indicator that the emergence of biotype 3 is a recent evolutionary event (14,16,36).
We identified independent risk factors for death in our group, including bacteremia, altered immune status, and history of ischemic heart disease. Septic shock was also found to be a strong predictor of death; however, septic shock may also be an outcome variable. Nonetheless, prior reports have also identified septic shock or hypotension as important risk factors for death (8,23). A recent large study from Taiwan (29) found that treatment with third-generation cephalosporins combined with tetracycline was an independent predictor of lower death rates in a subgroup of patients with hemorrhagic bullous necrotic cutaneous lesions. We did not demonstrate any correlation between a specific antimicrobial drug regimen and death rate in our study population. The predisposing diseases that were associated with death in previous reports were liver disease and neutropenia (8,23). Ischemic heart disease was not previously recognized as a classic predisposing factor predicting death; however, in the Vibrio-associated wound infections after Hurricane Katrina in Louisiana, USA, ischemic heart disease occurred in 7 of 13 patients (54%) with more severe illness (25).
Before the introduction of V. vulnificus biotype 3 into the fish aquaculture no infections were reported in Israel (37). Also, no reports have been made of V. vulnificus infection acquired through marine activities in the Mediterranean Sea. Almost the entire impact of V. vulnificus infections in Israel is associated with the freshwater fish industry. In response to the new threat, the Israeli Ministry of Health has issued regulations forbidding the selling of live, uncleaned tilapia (11,38). Fish stores and fishpond workers have been instructed to use protective gloves when handling fish, to keep fresh fish packed in ice, and to prevent direct contact between buyers and live fish. The public has been instructed accordingly. Apparently, compliance with these regulations is not universal.
Our findings outline the substantial effects of V. vulnificus illnesses in Israel and support a call for more strict regulations of fresh fish marketing as well as public education. Research efforts should focus on how V. vulnificus has penetrated the freshwater aquaculture in Israel and ways in which this trend can be reversed.
Dr Zaidenstein is an infectious diseases consultant at Assaf Harofeh Medical Center, Zerifin, Israel. Her recent research interest is the epidemiology of nosocomial and zoonotic infections.
References
- Neil MA, Carpenter CC. Other pathogenic vibrios. In: Mandell GL, Bennett JE, Dolin R, editors. Mandell, Douglas and Bennett’s principles and practice of infectious diseases. 5th ed. Philadelphia: Elsevier, Churchill Livingstone; 2005. p. 2545–47.
- Blake PA, Merson MH, Weaver RE, Hollis DG, Heublein PC. Disease caused by a marine Vibrio. Clinical characteristics and epidemiology. N Engl J Med. 1979;300:1–5.PubMedGoogle Scholar
- Morris JG Jr, Black RE. Cholera and other vibrioses in the United States. N Engl J Med. 1985;312:343–50.PubMedGoogle Scholar
- Hlady WG, Klontz KC. The epidemiology of Vibrio infections in Florida, 1981–1993. J Infect Dis. 1996;173:1176–83.PubMedGoogle Scholar
- Hlady WG, Mullen RC, Hopkin RS. Vibrio vulnificus from raw oysters. Leading cause of reported deaths from foodborne illness in Florida. J Fla Med Assoc. 1993;80:536–8.PubMedGoogle Scholar
- Wright AC, Simpson LM, Oliver JD. Role of iron in the pathogenesis of Vibrio vulnificus infections. Infect Immun. 1981;34:503–7.PubMedGoogle Scholar
- Haq SM, Dayal HH. Chronic liver disease and consumption of raw oysters: a potentially lethal combination—a review of Vibrio vulnificus septicemia. Am J Gastroenterol. 2005;100:1195–9. DOIPubMedGoogle Scholar
- Shapiro RL, Altekruse S, Hutwagner L, Bishop R, Hammond R, Wilson S, The role of Gulf Coast oysters harvested in warmer months in Vibrio vulnificus infections in the United States, 1988–1996. Vibrio Working Group. J Infect Dis. 1998;178:752–9.PubMedGoogle Scholar
- Strom MS, Paranjpye RN. Epidemiology and pathogenesis of Vibrio vulnificus. Microbes Infect. 2000;2:177–88. DOIPubMedGoogle Scholar
- Morris JG Jr. Cholera and other types of vibriosis: a story of human pandemics and oysters on the half shell. Clin Infect Dis. 2003;37:272–80. DOIPubMedGoogle Scholar
- Bisharat N, Agmon V, Finkelstein R, Raz R, Ben-Dror G, Lerner L, Clinical, epidemiological, and microbiological features of Vibrio vulnificus biogroup 3 causing outbreaks of wound infection and bacteraemia in Israel. Israel Vibrio Study Group. Lancet. 1999;354:1421–4. DOIPubMedGoogle Scholar
- Colodner R, Raz R, Meir I, Lazarovich T, Lerner L, Kopelowitz J, Identification of the emerging pathogen Vibrio vulnificus biotype 3 by commercially available phenotypic methods. J Clin Microbiol. 2004;42:4137–40. DOIPubMedGoogle Scholar
- Gutacker M, Conza N, Benagli C, Pedroli A, Bernasconi MV, Permin L, Population genetics of Vibrio vulnificus: identification of two divisions and a distinct eel-pathogenic clone. Appl Environ Microbiol. 2003;69:3203–12. DOIPubMedGoogle Scholar
- Bisharat N, Cohen DI, Harding RM, Falush D, Crook DW, Peto T, Hybrid Vibrio vulnificus. Emerg Infect Dis. 2005;11:30–5.PubMedGoogle Scholar
- Bisharat N, Cohen DI, Maiden MC, Crook DW, Peto T, Harding RM. The evolution of genetic structure in the marine pathogen, Vibrio vulnificus. Infect Genet Evol. 2007;7:685–93. DOIPubMedGoogle Scholar
- Bisharat N, Amaro C, Fouz B, Llorens A, Cohen DI. Serological and molecular characteristics of Vibrio vulnificus biotype 3: evidence for high clonality. Microbiology. 2007;153:847–56. DOIPubMedGoogle Scholar
- Bisharat N. Vibrio vulnificus infections can be avoided. Isr Med Assoc J. 2002;4:631–3.PubMedGoogle Scholar
- Cooper KL, Luey CK, Bird M, Terajima J, Nair GB, Kam KM, Development and validation of a PulseNet standardized pulsed-field gel electrophoresis protocol for subtyping of Vibrio cholerae. Foodborne Pathog Dis. 2006;3:51–8. DOIPubMedGoogle Scholar
- Romling U, Tummler B. Achieving 100% typeability of Pseudomonas aeruginosa by pulsed-field gel electrophoresis. J Clin Microbiol. 2000;38:464–5.PubMedGoogle Scholar
- Hunter SB, Vauterin P, Lambert-Fair MA, Van Duyne MS, Kubota K, Graves L, Establishment of a universal size standard strain for use with the PulseNet standardized pulsed-field gel electrophoresis protocols: converting the national databases to the new size standard. J Clin Microbiol. 2005;43:1045–50. DOIPubMedGoogle Scholar
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Eighteenth Informational Supplement. CLSI Document M100-S18. Wayne (PA): The Institute; 2008.
- Hsueh PR, Lin CY, Tang HJ, Lee HC, Liu JW, Liu YC, Vibrio vulnificus in Taiwan. Emerg Infect Dis. 2004;10:1363–8.PubMedGoogle Scholar
- Klontz KC, Lieb S, Schreiber M, Janowski HT, Baldy LM, Gunn RA. Syndromes of Vibrio vulnificus infections. Clinical and epidemiologic features in Florida cases, 1981–1987. Ann Intern Med. 1988;109:318–23.PubMedGoogle Scholar
- Chuang YC, Yuan CY, Liu CY, Lan CK, Huang AH. Vibrio vulnificus infection in Taiwan: report of 28 cases and review of clinical manifestations and treatment. Clin Infect Dis. 1992;15:271–6.PubMedGoogle Scholar
- Centers for Disease Control and Prevention. Vibrio illnesses after Hurricane Katrina—multiple states, August–September 2005. MMWR Morb Mortal Wkly Rep. 2005;54:928–31.PubMedGoogle Scholar
- Merkel SM, Alexander S, Zufall E, Oliver JD, Huet-Hudson YM. Essential role for estrogen in protection against Vibrio vulnificus-induced endotoxic shock. Infect Immun. 2001;69:6119–22. DOIPubMedGoogle Scholar
- Dechet AM, Yu PA, Koram N, Painter J. Non-foodborne Vibrio infections: an important cause of morbidity and mortality in the United States, 1997–2006. Clin Infect Dis. 2008;46:970–6. DOIPubMedGoogle Scholar
- Chung PH, Chuang SK, Tsang T, Wai-man L, Yung R, Lo J. Cutaneous injury and Vibrio vulnificus infection. Emerg Infect Dis. 2006;12:1302–3.PubMedGoogle Scholar
- Liu JW, Lee IK, Tang HJ, Ko WC, Lee HC, Liu YC, Prognostic factors and antibiotics in Vibrio vulnificus septicemia. Arch Intern Med. 2006;166:2117–23. DOIPubMedGoogle Scholar
- Dalsgaard I, Hoi L, Siebeling RJ, Dalsgaard A. Indole-positive Vibrio vulnificus isolated from disease outbreaks on a Danish eel farm. Dis Aquat Organ. 1999;35:187–94. DOIPubMedGoogle Scholar
- Calif E, Pick N, Dreyfuss U, Stahl S. Upper extremity infections following common carp fish (Cyprinus carpio) handling. J Hand Surg [Br]. 2002;27:78–82. DOIPubMedGoogle Scholar
- Morris JG Jr, Tenney JH, Drusano GL. In vitro susceptibility of pathogenic Vibrio species to norfloxacin and six other antimicrobial agents. Antimicrob Agents Chemother. 1985;28:442–5.PubMedGoogle Scholar
- Han F, Walker RD, Janes ME, Prinyawiwatkul W, Ge B. Antimicrobial susceptibilities of Vibrio parahaemolyticus and Vibrio vulnificus isolates from Louisiana Gulf and retail raw oysters. Appl Environ Microbiol. 2007;73:7096–8. DOIPubMedGoogle Scholar
- Wong HC, Chen SY, Chen MY, Oliver JD, Hor LI, Tsai WC. Pulsed-field gel electrophoresis analysis of Vibrio vulnificus strains isolated from Taiwan and the United States. Appl Environ Microbiol. 2004;70:5153–8. DOIPubMedGoogle Scholar
- Gonzalez-Escalona N, Whitney B, Jaykus LA, DePaola A. Comparison of direct genome restriction enzyme analysis and pulsed-field gel electrophoresis for typing of Vibrio vulnificus and their correspondence with multilocus sequence typing data. Appl Environ Microbiol. 2007;73:7494–500. DOIPubMedGoogle Scholar
- Broza YY, Danin-Poleg Y, Lerner L, Broza M, Kashi Y. Vibrio vulnificus typing based on simple sequence repeats: insights into the biotype 3 group. J Clin Microbiol. 2007;45:2951–9. DOIPubMedGoogle Scholar
- Paz S, Bisharat N, Paz E, Kidar O, Cohen D. Climate change and the emergence of Vibrio vulnificus disease in Israel. Environ Res. 2007;103:390–6. DOIPubMedGoogle Scholar
- Israel Ministry of Health Food Control Administration. Sho-2001. Announcement regarding handling of fresh fish. Tel Aviv (Israel): 1997 May 28.
Figures
Tables
Follow Up
CME FOLLOW-UP
Earning Medscape CME Credit
To obtain credit, you should first read the journal article. After reading the article, you should be able to answer the following, related, multiple-choice questions. To complete the questions and earn continuing medical education (CME) credit, please go to http://www.medscape.com/cme/eid. Credit cannot be obtained for tests completed on paper, although you may use the worksheet below to keep a record of your answers. You must be a registered user on Medscape.com. If you are not registered on Medscape.com, please click on the New Users: Free Registration link on the left hand side of the website to register. Only one answer is correct for each question. Once you successfully answer all post-test questions you will be able to view and/or print your certificate. For questions regarding the content of this activity, contact the accredited provider, CME@medscape.net. For technical assistance, contact CME@webmd.net. American Medical Association's Physician's Recognition Award (AMA PRA) credits are accepted in the US as evidence of participation in CME activities. For further information on this award, please refer to http://www.ama-assn.org/ama/pub/category/2922.html. The AMA has determined that physicians not licensed in the US who participate in this Medscape CME activity are eligible for AMA PRA Category 1 Credits™. Through agreements that the AMA has made with agencies in some countries, AMA PRA credit is acceptable as evidence of participation in CME activities. If you are not licensed in the US and want to obtain an AMA PRA CME credit, please complete the questions online, print the certificate and present it to your national medical association.
Medscape CME Questions
1. Which of the following is least likely to be a predisposing factor for infections caused by Vibrio vulnificus biotypes 1 and 2?
A. Anemia
B. Organ transplantation
C. HIV infection
D. Chronic liver disease
2. Illness caused by V. vulnificus biotype 3 is best distinguished from that caused by biotypes 1 and 2 by which of the following?
A. Method of transmission
B. Genetic features
C. Geographic distribution
D. All of the above
3. Which of the following is the primary fish involved in V. vulnificus biotype 3 infection?
A. Gray mullet
B. Tilapia
C. Salmon
D. Sea bass
4. Which of the following best describes the mortality associated with V. vulnificus biotype 3 infection as reported in Israel?
A. 1.0%
B. 2.3%
C. 5.2%
D. 7.5%
5. Which of the following is least likely to be an independent predictor of mortality for V. vulnificus biotype 3 infection?
A. Bacteremia
B. Ischemic heart disease
C. Sex
D. Altered immune status
Activity Evaluation
| 1. The activity supported the learning objectives. | ||||
| Strongly Disagree |
Strongly Agree
|
|||
| 1 |
2
|
3
|
4
|
5
|
| 2. The material was organized clearly for learning to occur. | ||||
| Strongly Disagree |
Strongly Agree
|
|||
| 1 |
2
|
3
|
4
|
5
|
| 3. The content learned from this activity will impact my practice. | ||||
| Strongly Disagree |
Strongly Agree
|
|||
| 1 |
2
|
3
|
4
|
5
|
| 4. The activity was presented objectively and free of commercial bias. | ||||
| Strongly Disagree |
Strongly Agree
|
|||
| 1 |
2
|
3
|
4
|
5
|
Related Links
Table of Contents – Volume 14, Number 12—December 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Miriam Weinberger, Infectious Diseases Unit, Assaf Harofeh Medical Center, Zerifin, Israel 70300
Top