Volume 22, Number 7—July 2016
Dispatch
Expanding Distribution of Lethal Amphibian Fungus Batrachochytrium salamandrivorans in Europe
Cite This Article
Citation for Media
Abstract
Emerging fungal diseases can drive amphibian species to local extinction. During 2010–2016, we examined 1,921 urodeles in 3 European countries. Presence of the chytrid fungus Batrachochytrium salamandrivorans at new locations and in urodeles of different species expands the known geographic and host range of the fungus and underpins its imminent threat to biodiversity.
Amphibians provide an iconic example of disease-driven global loss in biodiversity. The recently described chytrid fungus Batrachochytrium salamandrivorans (Bsal) is an emerging pathogen that is driving amphibian populations to local extinction (1,2). This highly pathogenic fungus causes a lethal skin disease that has so far been restricted to urodele amphibians (newts and salamanders); the fungus was most likely introduced from East Asia into Europe via the pet trade (2). In Europe, Bsal infection has led to dramatic declines of fire salamander (Salamandra salamandra) populations in the Netherlands and Belgium (2). Within 7 years after the supposed introduction of the fungus, a population in the Netherlands declined by 99.9% (3,4). In the United Kingdom and Germany, Bsal has been detected in captive salamanders and newts (5,6). Infection trials suggest that Bsal represents an unprecedented threat to diversity of Western Palearctic urodeles (2); nevertheless, reports of deaths among salamanders and newts in their naturalized ranges have been restricted to a few populations of a single salamander species in the southern Netherlands and adjacent Belgium (1,3). Using data from field surveillance, we examined the hosts and the geographic range of Bsal in Europe.
Figure
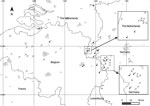
Figure. Study sites for collection of amphibians in Western Europe, 2010–2016. Numbers correspond to field sites at which amphibians were collected and examined for Batrachochytrium salamandrivorans (Bsal) (Technical Appendix)....
During 2010–2016, we collected samples of free-living populations of newts and salamanders from 48 sites in the Netherlands, Belgium, and adjacent regions of the Eifel region in Germany (near the border with the Netherlands and Belgium) (Figure; Technical Appendix Table 1). Site selection was based on reported amphibian deaths, apparent negative amphibian population trends, preventive Bsal surveillance in susceptible populations, or geographic proximity to known outbreak sites. Samples were also collected at 6 additional sites in Germany and 1 in the Netherlands, which were located >100 km from the nearest known outbreak (Technical Appendix Table 2). Sampling was conducted by swabbing skin (7,8) of live animals and collecting skin samples from dead animals. All samples were kept frozen at −20°C until they were analyzed for the presence of Bsal DNA via real-time PCR, as described (9).
Across all 55 sites, we tested 1,019 fire salamanders (43 dead, 976 skin swab samples); at site 14, skin swab samples instead of tissue samples were collected from 16 dead salamanders. We also collected samples from 474 alpine newts (Ichthyosaura alpestris; 18 dead, 456 skin swab samples), 239 smooth newts (Lissotriton vulgaris; 2 dead, 237 skin swab samples), 80 palmate newts (Lissotriton helveticus; all skin swab samples), 79 crested newts (Triturus cristatus; all skin swab samples), and 30 Italian crested newts (Triturus carnifex; all skin swab samples). To obtain a Bayesian 95% credible interval for prevalence (online Technical Appendix), we used the computational methods of Lötters et al. (10). We ran 3 parallel Markov chains with 20,000 iterations each and discarded the first 5,000 iterations as burn-in; chains were not thinned.
Bsal was found at 14 of the 55 sites; infected amphibians were fire salamanders, alpine newts, and smooth newts. Our results demonstrate that the range of Bsal distribution may be up to ≈10,000 km2 (measured as the surface of a minimum convex polygon encompassing the outermost points) across Germany, Belgium, and the Netherlands (Figure). The presence of Bsal in wild alpine newts and smooth newts shows distinct expansion of the known host range in the wild (Technical Appendix Table 1). Furthermore, we document that Bsal is present in natural fire salamander populations in Germany (confined to the Eifel region). At some sites, because of our sample sizes, the upper limit of the 95% credible interval for Bsal prevalence was as high as 0.7; therefore, we may have failed to detect Bsal at these sites (Technical Appendix Table 1). In addition, the fungus may have been present at several sites before first detection. For example, Bsal was detected at site 4, where population-monitoring efforts in the years before detection (2000–2013) showed declines in 4 newt species (http://www.ravon.nl/EID_SI_Spitzen_et_al_2016). However, because no samples were collected before 2015, we have no evidence for a causal relationship between the presence of Bsal and the declines. We have also recorded the presence of Bsal in populations with no evidence of population change so far, such as the incidental findings of dead Bsal-positive newts in fyke nets at sites 5 and 11, and the incidental findings of dead Bsal-positive fire salamanders at sites 12 and 14. Clinical signs of mycosis, such as lethargy and skin shedding (1), were observed at some Bsal-positive sites (1, 2, 7, 8, 14) but not at others.
Our study provides evidence that Bsal among wild amphibians in Europe is more widely distributed and affects a wider host range than previously known, which can either indicate recent spread of the fungus or point to historically infected sites that hitherto remained undetected. The presence of Bsal in wild populations can easily remain unnoticed because the lesions develop only near the final stage of the disease (1). This information is crucial for the design of field surveys for Bsal surveillance. Our data might be used to inform a management strategy and to implement the recommendation of the Bern Convention (11) to halt the spread of Bsal in Europe. Research to search for molecular evidence that the outbreak locations are connected is under way. Chytrid disease dynamics are affected by multiple factors (e.g., temperature regimes [1]), and yet undetermined environmental determinants might be essential for disease outbreaks (12). Untangling these factors, as well as the modes of Bsal spread and its geographic distribution, are points for further research to fully map the problem and identify populations and species at risk.
Ms. Spitzen-van der Sluijs is a senior project manager at Reptile, Amphibian & Fish Conservation Netherlands and a PhD candidate at Ghent University. Her work focuses on the effects of emerging infectious diseases on native amphibian population dynamics.
Acknowledgments
We thank A. Geiger, K. Fischer, W. Bosman, M. Haffmann, J. Janse, J. Munro, L. Paulssen, E. Schotmans, and S. van Baren for their invaluable help and support to this project. We also thank all volunteers from Hyla, Natagora and Reptile, Amphibian & Fish Conservation Netherlands for collecting amphibians.
This study was supported by the Deutsche Bundesstiftung Umwelt, Dutch Ministry of Economic Affairs, Flemish Agency for Nature and Forests, Land Nordrhein Westfalen, Province of Limburg, StädteRegion Aachen, Stiftung Artenschutz, Trier University Research Assistantship, Verband der Zoologischen Gärten, Zoo Landau, and a PhD fellowship of the Research Foundation–Flanders.
References
- Martel A, Spitzen-van der Sluijs A, Blooi M, Bert W, Ducatelle R, Fisher MC, Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proc Natl Acad Sci U S A. 2013;110:15325–9 .DOIPubMedGoogle Scholar
- Martel A, Blooi M, Adriaensen C, Van Rooij P, Beukema WM, Fisher MC, Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science. 2014;346:630–1 . DOIPubMedGoogle Scholar
- Spitzen-van der Sluijs A, Spikmans F, Bosman W, De Zeeuw M, Van der Meij T, Goverse E, Rapid enigmatic decline drives the fire salamander (Salamandra salamandra) to the edge of extinction in the Netherlands. Amphib-Reptil. 2013;34:233–9 .DOIGoogle Scholar
- Goverse E, De Zeeuw M. Schubben & slijm: resultaten meetnet amfibieën 2014 [cited 2016 Mar 21]. http://www.ravon.nl/Portals/0/PDF3/schubbenslijm26.pdf
- Cunningham AA, Beckmann K, Perkins M, Fitzpatrick L, Cromie R, Redbond J, Emerging disease in UK amphibians. Vet Rec. 2015;176:468. DOIPubMedGoogle Scholar
- Sabino-Pinto J, Bletz M, Hendrix R, Perl RGB, Martel A, Pasmans F, First evidence of Batrachochytrium salamandrivorans from a captive salamander population in Germany. Amphib-Reptil. 2015;36:411–6. DOIGoogle Scholar
- Hyatt AD, Boyle DG, Olsen V, Boyle DB, Berger L, Obendorf D, Diagnostic assays and sampling protocols for the detection of Batrachochytrium dendrobatidis. Dis Aquat Organ. 2007;73:175–92. DOIPubMedGoogle Scholar
- Van Rooij P, Martel A, Nerz J, Voitel S, Van Immerseel F, Haesebrouck F, Detection of Batrachochytrium dendrobatidis in Mexican bolitoglossine salamanders using an optimal sampling protocol. EcoHealth. 2011;8:237–43. DOIPubMedGoogle Scholar
- Blooi M, Pasmans F, Longcore JE, Spitzen-van der Sluijs A, Vercammen F, Martel A. Duplex real-time PCR for rapid simultaneous detection of Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans in amphibian samples. J Clin Microbiol. 2013;51:4173–7. DOIPubMedGoogle Scholar
- Lötters S, Kielgast J, Sztatecsny M, Wagner N, Schulte U, Werner P, Absence of infection with the amphibian chytrid fungus in the terrestrial alpine salamander, Salamandra atra. Salamandra. 2012;48:58–62.
- Council of Europe. Convention on the Conservation of European Wildlife and Natural Habitats, 35th meeting; 2015 Dec 1–4; Strasbourg, France. Recommendation no. 176 (2015) on the prevention and control of the Batrachochytrium salamandrivorans chytrid fungus [cited 2016 Mar 21]. https://wcd.coe.int/com.instranet.InstraServlet?command=com.instranet.CmdBlobGet&InstranetImage=2859613&SecMode=1&DocId=2339082&Usage=2
- Van Rooij P, Martel A, Haesenbrouck F, Pasmans F. Amphibian chytridiomycosis: a review with focus on fungus-host interactions. Vet Res. 2015;46:137.PubMedGoogle Scholar
Figure
Cite This ArticleTable of Contents – Volume 22, Number 7—July 2016
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Stefan Lötters, Trier University, Biogeography, 54286 Trier, Germany
Top