Volume 23, Number 3—March 2017
Dispatch
Pneumonic Plague Transmission, Moramanga, Madagascar, 2015
Cite This Article
Citation for Media
Abstract
During a pneumonic plague outbreak in Moramanga, Madagascar, we identified 4 confirmed, 1 presumptive, and 9 suspected plague case-patients. Human-to-human transmission among close contacts was high (reproductive number 1.44) and the case fatality rate was 71%. Phylogenetic analysis showed that the Yersinia pestis isolates belonged to group q3, different from the previous outbreak.
Plague, caused by the bacterium Yersinia pestis, is a fleaborne disease responsible for the death of tens of millions of persons throughout history (1,2). The bacterium spread to Madagascar in 1898 through trade routes and became endemic in the country’s central highlands (3). Despite considerable control efforts, Y. pestis remains a public health threat in Madagascar (4–6). For the past 2 decades, Madagascar reported the highest number of human infections in the world (7).
Bubonic plague is the most common disease form resulting from the bite of an infected flea. As bubonic plague worsens, it can progress to pneumonic plague, in which persons expel infectious, aerosolized respiratory droplets. During pneumonic plague outbreaks, person-to-person transmission facilitates the spread from the initial infected person to family members and the wider community (5,8). Pneumonic plague is rare but poses a substantial danger in Madagascar because of the country’s relatively weak healthcare system (4,6) and widespread traditional burial methods involving interaction with corpses. Understanding how pneumonic plague outbreaks spread is vital for managing disease prevention and educating the population. We investigated the transmission chain of a pneumonic plague outbreak that occurred in Madagascar outside the normal plague season (October–March) (9) in a remote area that had been free of human plague for 13 years.
Figure 1

Figure 1. Course of pneumonic plague outbreak in Moramanga, Madagascar, August 17–September 1, 2015 (N = 14). Each line corresponds to a case-patient and describes disease outcomes. The vertical dashed line denotes when...
Over a 10-day period, 14 persons became ill with pneumonia and fever. The first case-patient (case-patient 1) displayed symptoms on August 17, 2015, while in a remote area in Antsahatsianarina hamlet, Tsiazompody village, Ampasipotsy Gara commune, within the Moramanga district of Madagascar (Technical Appendix Table). Case-patient 1 was a 22-year-old man who had traveled 1 week before disease onset. After returning home, he experienced chest pain, fever, and cough. Two days later, his condition deteriorated and, while relatives and neighbors assisted him to the nearest health center, he died near Beravina, Madagascar. He was buried near Beravina in a traditional manner with a 2-night wake, exposing the family and community to the pathogen and initiating a chain of transmission. On August 22 and 23, plague symptoms developed in members of case-patient 1’s immediate family (N = 2), his extended family (N = 6), and the community (N = 3). Of these, 4 died, and 7 self-referred to the Moramanga district hospital, where 4 died upon arrival, and 3 survived after a 6- to 9-day course of antimicrobial drug treatment. On August 25, a second chain of transmission (N = 2) was identified (Figure 1). One case-patient was from the outbreak epicenter, and the other was a suspected nosocomial transmission; no connection to the other cases could be found except occupying the same hospital space as some of the earlier case-patients. Transmission ceased with these last 2 case-patients.
Figure 2
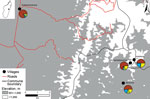
Figure 2. Map of pneumonic plague outbreak (n = 14) in the commune of Ampasipotsy Gara in Moramanga, Madagascar, 2015. The index case-patient (case-patient 1) was infected with Yersinia pestis at Antsahatsianarina and...
Before illness onset, case-patients 2–14 had social or spatial contact with a symptomatic person who could be traced back to case-patient 1. Two of the deceased case-patients (2 and 10) were visitors to Antsahatsianarina who had returned home to their respective communities Ambatoharanana and Ambilona (Figure 2), potentially spreading the bacterium to others. To investigate the extent of pathogen spread, we identified 123 case-patient contacts in 4 communities connected with case-patient 1 (Technical Appendix Table). On August 24, 2015, an outbreak investigation protocol was applied by the Institut Pasteur de Madagascar and the Malagasy Ministry of Health, whose ethics committee approved the study (068-MSANP/CE). Verbal consent was obtained from 71 contacts to test their serum for plague antibody with a capsular antigen fraction 1 (F1) IgG ELISA (10,11); 7/20 (35%) contacts from Antsahatsianarina, 12/20 (60%) from Beravina (the burial site of case-patient 1), 9/10 (90%) from Ambilona, and 7/21 (33%) from Ambatoharanana were seropositive (Figure 2; Technical Appendix Table). The 35 contacts positive for F1-specific antibody were given chemoprophylaxis, and their infections remained subclinical.
The 14 case-patients who had sought medical care for pneumonia and fever had clinical signs and symptoms consistent with those for plague. To confirm Y. pestis infection, we performed bacteria culture on sputum or organ puncture (12) and serologic testing of serum samples (10). According to the international standards definition (13), 4/5 patients (1 deceased) were confirmed positive for Y. pestis (2 by culture and 2 by seroconversion) and 1/5 was presumptive for Y. pestis infection. No samples were collected from the other 9 (all deceased) persons, who were thus considered suspected plague case-patients (13).
More men (n = 10) than women (n = 4) were infected. Median patient age was 22.5 (range 15–80) years, and the overall case-fatality rate was 71%. The most common signs and symptoms were fever, dyspnea, chest pain, and cough (100% of patients); blood-stained sputum (93%); chills (86%); and headache (71%). The average duration from onset of full-blown disease to death was 1.9 days, and the average infectious period was 3.5 days. Basic reproductive number (R0) was 1.44, and the transmission rate was 0.41 susceptibles/day (14).
To investigate the possible outbreak origin, we trapped 100 rats and mice over a period of 3 nights at Antsahatsianarina, Beravina, and Ambilona. Rodent spleens were sampled in accordance with the directive 2010/63/EU of the European Parliament (http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:EN:PDF) and tested by using an F1 antigen rapid diagnostic test (3). A total of 22 (22%) rats, but no mice, from the 3 communities tested positive for plague antigen (Table).
The lack of human plague activity in this area for the past 13 years suggests that a reservoir species, such as rodents or fleas, transmitted bubonic plague to case-patient 1 and that the disease later progressed into the pneumonic form. Relatives of case-patient 1 indicated that moribund rodents were present near his home 2 weeks before he became ill. The family who hosted case-patient 1 during his travels (1 week before his disease onset) reported no disease present in their household. Taken together, the epidemiologic data suggest that case-patient 1’s hamlet (Antsahatsianarina) was the probable outbreak epicenter.
Using published single-nucleotide polymorphisms (15), we assigned isolates from 2 case-patients and 1 recently sampled rat to the Y. pestis q3 phylogenetic subgroup within group I (node k). The matched genetic grouping between the 2 human samples is consistent with human-to-human transmission. The matched genetic grouping among all 3 samples is consistent with Y. pestis in the initial case-patient originating from the environment rather than another human. We compared these phylogenetic data with data from 4 archived isolates (human and rodent) obtained in 2000, 2002, and 2003 from the same area. The archived isolates were also assigned to group I (node k), but unlike the isolates from this study, they were assigned to the q2 subgroup (15). This finding indicates that the recent outbreak did not arise from the same phylogenetic groups responsible for past outbreaks and illustrates how outbreaks in different years are probably conferred by Y. pestis from different environmental sources. As of January 2017, 4 lineages (q1, q2, q3, and q4) have been recorded in Moramanga, suggesting multiple genotypes are persisting within this region (15).
Despite Madagascar having an effective surveillance system, plague control remains a public health challenge. Pneumonic plague is rare but persists as a threat in Madagascar, where poor healthcare systems and traditional burial practices promote these outbreaks. Our findings provide additional understanding of pneumonic plague transmission patterns, argues for continued public education, and informs authorities about effective outbreak response practices.
Dr. Ramasindrazana is an ecologist–mammalogist in the Plague Unit at Institut Pasteur de Madagascar. He conducts ecological and epidemiological surveys on small mammals to investigate host-parasite relationships.
Acknowledgments
We are grateful to the staff of Moramanga District Hospital and the Ministry of Health for assistance in different aspects during the outbreak response, to the staff of the Plague Central Laboratory–Institut Pasteur de Madagascar for technical assistance, to Dave Wagner for editing this manuscript, and to Anthonio Rakotoarison for study site mapping.
This study was supported by the Institut Pasteur de Madagascar.
References
- Perry RD, Fetherston JD. Yersinia pestis—etiologic agent of plague. Clin Microbiol Rev. 1997;10:35–66.PubMedGoogle Scholar
- Keim PS, Wagner DM. Humans and evolutionary and ecological forces shaped the phylogeography of recently emerged diseases. Nat Rev Microbiol. 2009;7:813–21. DOIPubMedGoogle Scholar
- Chanteau S, Rahalison L, Ralafiarisoa L, Foulon J, Ratsitorahina M, Ratsifasoamanana L, et al. Development and testing of a rapid diagnostic test for bubonic and pneumonic plague. Lancet. 2003;361:211–6. DOIPubMedGoogle Scholar
- Andrianaivoarimanana V, Kreppel K, Elissa N, Duplantier JM, Carniel E, Rajerison M, et al. Understanding the persistence of plague foci in Madagascar. PLoS Negl Trop Dis. 2013;7:e2382. DOIPubMedGoogle Scholar
- Richard V, Riehm JM, Herindrainy P, Soanandrasana R, Ratsitoharina M, Rakotomanana F, et al. Pneumonic plague outbreak, Northern Madagascar, 2011. Emerg Infect Dis. 2015;21:8–15. DOIPubMedGoogle Scholar
- Bertherat EG; World Health Organization. Plague in Madagascar:overview of the 2014-2015 epidemic season. Wkly Epidemiol Rec. 2015;90:250–2.PubMedGoogle Scholar
- World Health Organization. Plague around the world, 2010–2015. Wkly Epidemiol Rec. 2016;91:89–93.PubMedGoogle Scholar
- Begier EM, Asiki G, Anywaine Z, Yockey B, Schriefer ME, Aleti P, et al. Pneumonic plague cluster, Uganda, 2004. Emerg Infect Dis. 2006;12:460–7. DOIPubMedGoogle Scholar
- Migliani R, Chanteau S, Rahalison L, Ratsitorahina M, Boutin JP, Ratsifasoamanana L, et al. Epidemiological trends for human plague in Madagascar during the second half of the 20th century: a survey of 20,900 notified cases. Trop Med Int Health. 2006;11:1228–37. DOIPubMedGoogle Scholar
- Rasoamanana B, Leroy F, Boisier P, Rasolomaharo M, Buchy P, Carniel E, et al. Field evaluation of an immunoglobulin G anti-F1 enzyme-linked immunosorbent assay for serodiagnosis of human plague in Madagascar. Clin Diagn Lab Immunol. 1997;4:587–91.PubMedGoogle Scholar
- Dromigny JA, Ralafiarisoa L, Raharimanana C, Randriananja N, Chanteau S. La sérologie anti-F1 chez la souris OF1, test complémentaire pour le diagnostic de la peste humaine. Arch Inst Pasteur Madagascar. 1998;64:18–20.
- Rasoamanana B, Rahalison L, Raharimanana C, Chanteau S. Comparison of Yersinia CIN agar and mouse inoculation assay for the diagnosis of plague. Trans R Soc Trop Med Hyg. 1996;90:651. DOIPubMedGoogle Scholar
- World Health Organization. International meeting on preventing and controlling plague: the old calamity still has a future. Wkly Epidemiol Rec. 2006;81:278–84.PubMedGoogle Scholar
- Gani R, Leach S. Epidemiologic determinants for modeling pneumonic plague outbreaks. Emerg Infect Dis. 2004;10:608–14. DOIPubMedGoogle Scholar
- Vogler AJ, Chan F, Wagner DM, Roumagnac P, Lee J, Nera R, et al. Phylogeography and molecular epidemiology of Yersinia pestis in Madagascar. PLoS Negl Trop Dis. 2011;5:e1319. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 23, Number 3—March 2017
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Minoarisoa Rajerison, Plague Unit, Institut Pasteur de Madagascar, BP 1274 Ambatofotsikely Antananarivo-101, Madagascar
Top