Volume 28, Number 5—May 2022
Dispatch
Intercontinental Movement of Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4 Virus to the United States, 2021
Cite This Article
Citation for Media
Abstract
We detected Eurasian-origin highly pathogenic avian influenza A(H5N1) virus belonging to the Gs/GD lineage, clade 2.3.4.4b, in wild waterfowl in 2 Atlantic coastal states in the United States. Bird banding data showed widespread movement of waterfowl within the Atlantic Flyway and between neighboring flyways and northern breeding grounds.
Influenza A viruses have a worldwide distribution, and wild birds are the primary wild reservoir. Many wild ducks in particular are often repeatedly exposed to and infected with these viruses (hereafter referred to as avian influenza viruses or AIV) with little to no sign of clinical disease (1), although highly pathogenic forms of the virus can sometimes cause illness and death in wild birds (2). Highly pathogenic lineage viruses identified in 1996 (A/goose/Guangdong/1/1996 [Gs/GD]) have repeatedly spilled over from poultry to wild birds, and eventual emergence of highly pathogenic AIV Gs/GD clade 2.3.4.4 has led to more persistent circulation of these viruses in wild birds and high numbers of illnesses and deaths in poultry on multiple continents (3).
One way to better understand AIV movement on the landscape or to identify routes of introduction of novel AIVs is through wild bird band-recovery data (4). These data have been collected as part of waterfowl management and conservation efforts in North America since the 1920s (5). Spatial locations of where birds are banded and later recovered are recorded and archived, providing data on wild bird movement. For waterfowl, recoveries primarily occur through banded birds being reported as part of hunter harvest activities.
Wild bird samples are routinely collected by the US Department of Agriculture, Animal Plant Health Inspection Service, Wildlife Services, National Wildlife Disease Program (National Wildlife Disease Program, US Fish and Wildlife Service permit no. MB124992 0) and screened for AIV in conjunction with the National Animal Health Laboratory Network and with the National Veterinary Services Laboratories (Ames, Iowa, USA) as part of a targeted AIV surveillance program in wild birds (6). Samples analyzed in this investigation came from routine wild bird surveillance activities in the US Atlantic Flyway and were primarily obtained from hunter harvest activities, live-trapping, and bird banding operations. These surveillance data, combined with bird band-recovery movement data, can shed light on AIV occurrence on the landscape, and findings in wild birds can act as an early warning system for spillover risk to poultry and humans (6).
For these analyses, we initially screened wild bird samples by using an influenza matrix gene real-time, reverse transcription PCR. We then tested matrix gene presumptive positive samples by using H5 and H7 subtype-specific, real-time reverse transcription PCRs. Influenza A virus RNA from wild bird samples was amplified as described (7). After amplificationwas completed, we generated cDNA libraries for MiSeq by using the Nextera XT DNA Sample Preparation Kit (Illumina, https://www.illumina.com) and the 500 cycle MiSeq Reagent Kit v2 (Illumina) according to manufacturer instructions. We performed de novo and directed assembly of genome sequences by using IRMA version 0.6.7 (8), followed by visual verification in DNAStar SeqMan version 14 (https://www.dnastar.com). For phylogenetic analysis, we downloaded sequences from GISAID (https://www.gisaid.org) and aligned in Geneious 11.1.5 by using MAFFT (https://www.geneious.com), then generated trees by using RAxML (https://cme.h-its.org).
We queried North American Bird Banding Program data (5) to find all records from 1960–2021 for 11 dabbling duck species targeted for wild bird surveillance. These species were American black duck (Anas rubripes), American green-winged teal (Anas crecca carolinensis), American wigeon (Mareca americana), blue-winged teal (Spatula discors), cinnamon teal (Spatula cyanoptera), gadwall (Mareca strepera), mallard (Anas platyrhynchos), mottled duck (Anas fulvigula), northern pintail (Anas acuta), northern shoveler (Spatula clypeata), and wood duck (Aix sponsa). We then limited records for these species to only include birds that were either banded or encountered in North Carolina or South Carolina, USA, and >1 other state or province.
Figure 1

Figure 1. Maximum-likelihood phylogenic analysis of the hemagglutinin gene segment of the first sequenced set of wild bird isolates of highly pathogenic avian influenza A(H5N1) clade 2.3.4.4 virus, United States, 2021. Red...
As part of these routine surveillance efforts, we detected Gs/GD lineage clade 2.3.4.4b H5N1 highly pathogenic AIVs in multiple wild birds sampled in North Carolina and South Carolina during December 2021 and January 2022 (Figure 1). Genetic analyses showed that all virus segments were of Eurasian origin (99.7%–99.8% similar; Appendix) and have high identity with December 2021 AIV H5N1 findings in Newfoundland, Canada (Figure 1) (9).
A sample was collected on December 30, 2021 from an American wigeon in Colleton County, South Carolina [A/American_wigeon/South_Carolina/AH0195145/2021(H5N1), GISAID accession no. EPI_ISL_9869760]. Immediately after this finding, there was an additional wild bird detection in South Carolina [A/blue-winged_teal/South_Carolina/AH0195150/2021(H5N1), GISAID accession no. EPI_ISL_9876777] and detections in neighboring North Carolina (Figure 1). Another 291 detections in wild birds occurred within 2 months, indicating high susceptibility to infection with a novel virus along with continued transmission and dispersal (Table). All birds were apparently healthy live-trapped or hunter-obtained dabbling ducks (Appendix Table). North American lineage AIV was not found in any of these samples.
Figure 2
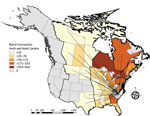
Figure 2. Dabbling duck movements to and from North Carolina and South Carolina, USA, to and from other states or provinces in study of highly pathogenic avian influenza A(H5N1) 2.3.4.4 virus, United...
Analysis of North American Bird Banding Program data showed broadscale movement of waterfowl throughout North America. Across 11 species of dabbling ducks targeted in surveillance sampling that were historically banded or encountered in North Carolina or South Carolina (and subsequently or previously banded or encountered in another state or province), a total of 64.7% of bird movements were within the Atlantic Flyway, 33.6% of analyzed species were encountered in the Atlantic and the Mississippi Flyways, and 1.7% were encountered in the Atlantic and Central Flyways (Figure 2).
Although there has been intense focus on intercontinental movement of highly pathogenic AIV from Asia to the North American Pacific Flyway (10), viral movement by the trans-Atlantic pathway has been less clear (11). Data reported here, in combination with the recent highly pathogenic AIV findings in Newfoundland, Canada (9), suggest that wild bird surveillance captured the introduction of a Eurasian-origin highly pathogenic AIV into wild birds by the Atlantic Flyway of the United States. The potential introduction pathway probably includes wild bird migratory routes from northern Europe that overlap Arctic regions of North America and then dispersal farther south into Canada and the United States (12).
Band recovery data showed that most dabbling ducks banded in the Atlantic Flyway are also recovered in the Atlantic Flyway, reinforcing the predominance of within flyway movement (13). However, data also show routine movement to other flyways, providing a potential mechanism of wider spread dispersal of the virus in North America.
In addition, sequence data indicate that these viruses cluster closely with viruses found in Western Europe during spring of 2021 (Figure 1; Appendix). If viruses were exchanged between North American and Eurasian waterfowl on northern breeding grounds during spring and summer 2021, and then carried south during fall of 2021, these viruses might already be in multiple locations in North America (Figure 2). Because wild bird surveillance has recently been limited to the Atlantic and Pacific Flyways, introductions into the Central or Mississippi Flyways might have gone undetected. Additional detections in wild birds suggest these clade 2.3.4.4b H5 viruses continue to be transmitted (Appendix Table), and further dispersal might be seen once waterfowl migrate to summer breeding areas.
Some findings of highly pathogenic AIVs in wild birds have been associated with repeated spillover of the viruses from domestic birds, which are where mutations to high pathogenicity primarily occur; however, in some cases, Gs/GD lineage viruses now appear to be maintained in wild bird populations (14). This potential adaptation of highly pathogenic AIV to wild birds highlights the need for continued wild bird surveillance. In addition, these findings demonstrate that targeted AIV surveillance in wild bird populations can detect newly introduced or emergent AIVs before spillover to domestic poultry. Advanced warnings from wild bird surveillance enable poultry producers to consider altering biosecurity in the face of increased AIV risk and also help inform zoonotic disease potential (15).
Dr. Bevins is a research scientist at the US Department of Agriculture National Wildlife Research Center, Fort Collins, CO. Her primary research interest is pathogen emergence from wild animals.
Acknowledgments
We thank Wildlife Services employees and collaborators at state wildlife agencies for contributing wildlife sampling expertise, hunters for participating in this large-scale effort, and staff at state and federal agency laboratories and at the Canadian Food Inspection Agency for providing contributions to this study.
This study was supported by the US Department of Agriculture, Animal Plant Health Inspection Service.
References
- Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. Evolution and ecology of influenza A viruses. Microbiol Rev. 1992;56:152–79. DOIPubMedGoogle Scholar
- Pohlmann A, Starick E, Harder T, Grund C, Höper D, Globig A, et al. Outbreaks among wild birds and domestic poultry caused by reassorted influenza A(H5N8) clade 2.3.4.4 viruses, Germany, 2016. Emerg Infect Dis. 2017;23:633–6. DOIPubMedGoogle Scholar
- Verhagen JH, Fouchier RAM, Lewis N. Highly pathogenic avian influenza viruses at the wild-domestic bird interface in Europe: future directions for research and surveillance. Viruses. 2021;13:212. DOIPubMedGoogle Scholar
- Franklin AB, Bevins SN, Ellis JW, Miller RS, Shriner SA, Root JJ, et al. Predicting the initial spread of novel Asian origin influenza A viruses in the continental USA by wild waterfowl. Transbound Emerg Dis. 2019;66:705–14. DOIPubMedGoogle Scholar
- Celis-Murillo A, Malorodova M, Nakash E. North American bird banding program dataset 1960–2021, retrieved 2021–07–25. US Geological Survey. 2021 [cited 2022 Mar 10]. https://www.sciencebase.gov/catalog/item/613f75d7d34e1449c5d35c77
- United States Interagency Working Group. Surveillance plan for highly pathogenic avian influenza in wild migratory birds in the United States. USDA Avian Influenza Disease. 2017 [cited 2022 Mar 10]. https://www.aphis.usda.gov/animal_health/downloads/animal_diseases/ai/2017-hpai-surveillance-plan.pdf
- Crossley BM, Rejmanek D, Baroch J, Stanton JB, Young KT, Killian ML, et al. Nanopore sequencing as a rapid tool for identification and pathotyping of avian influenza A viruses. J Vet Diagn Invest. 2021;33:253–60. DOIPubMedGoogle Scholar
- Shepard SS, Meno S, Bahl J, Wilson MM, Barnes J, Neuhaus E. Viral deep sequencing needs an adaptive approach: IRMA, the iterative refinement meta-assembler. BMC Genomics. 2016;17:708. DOIPubMedGoogle Scholar
- Avian influenza (04): Americas (Canada) wild bird. ProMed-mail [cited 2022 Mar 11]. https://www.promedmail.org, archive no. 20220103.8700643.
- Lee DH, Torchetti MK, Winker K, Ip HS, Song CS, Swayne DE. Intercontinental Spread of Asian-Origin H5N8 to North America through Beringia by Migratory Birds. J Virol. 2015;89:6521–4. DOIPubMedGoogle Scholar
- Makarova NV, Kaverin NV, Krauss S, Senne D, Webster RG. Transmission of Eurasian avian H2 influenza virus to shorebirds in North America. J Gen Virol. 1999;80:3167–71. DOIPubMedGoogle Scholar
- Dusek RJ, Hallgrimsson GT, Ip HS, Jónsson JE, Sreevatsan S, Nashold SW, et al. North Atlantic migratory bird flyways provide routes for intercontinental movement of avian influenza viruses. PLoS One. 2014;9:
e92075 . DOIPubMedGoogle Scholar - Li L, Bowman AS, DeLiberto TJ, Killian ML, Krauss S, Nolting JM, et al. Genetic evidence supports sporadic and independent introductions of subtype H5 low-pathogenic avian influenza A viruses from wild birds to domestic poultry in North America. J Virol. 2018;92:e00913–8. DOIPubMedGoogle Scholar
- Caliendo V, Leijten L, van de Bildt M, Germeraad E, Fouchier RAM, Beerens N, et al. Tropism of highly pathogenic avian influenza H5 viruses from the 2020/2021 epizootic in wild ducks and geese. Viruses. 2022;14:280. DOIPubMedGoogle Scholar
- Oliver I, Roberts J, Brown CS, Byrne AM, Mellon D, Hansen R, et al. A case of avian influenza A(H5N1) in England, January 2022. Euro Surveill. 2022;27:
2200061 . DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: March 18, 2022
1These authors contributed equally to this article.
Table of Contents – Volume 28, Number 5—May 2022
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Sarah N. Bevins, US Department of Agriculture National Wildlife Research Center, 4101 Laporte Ave, Fort Collins, CO, 80521, USA
Top