Volume 30, Number 6—June 2024
Dispatch
IMI-Type Carbapenemase-Producing Enterobacter cloacae Complex, France and Overseas Regions, 2012–2022
Cite This Article
Citation for Media
Abstract
We characterized a collection of IMI-like–producing Enterobacter spp. isolates (n = 112) in France. The main clone corresponded to IMI-1–producing sequence type 820 E. cloacae subspecies cloacae that was involved in an outbreak. Clinicians should be aware of potential antimicrobial resistance among these bacteria.
The Enterobacter cloacae complex (ECC) is highly diverse; its many species and subspecies can be distinguished by using phenotypic methods or matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Whole-genome sequencing enables the precise determination of the bacterial species inside this complex; 22 species, including 6 subspecies, have been assigned to the ECC. IMI and NmcA, which are Ambler class A carbapenemases conferring antimicrobial resistance, are typically associated with the ECC (1), but they are rarely reported in other bacterial species (2,3) despite a worldwide distribution.
A total of 24 NmcA/IMI-type variants have been identified in accordance with the Beta-Lactamase DataBase (http://www.bldb.eu) (4). The blaIMI/NmcA genes can be either chromosome or plasmid encoded; blaNmcA, blaIMI-1, blaIMI-4 and blaIMI-9 have been described as chromosome encoded (5–7). The chromosome-encoded blaIMI/NmcA genes are usually described into XerC/XerD recombinase-dependent integrative mobile elements (IMEX) called EcloIMEX elements. For all IMI producers, the genetic features showed an integration of EcloIMEX structures at the same position between setB and yeiP genes. For chromosomal variant, the blaIMI gene were mostly identified in E. cloacae subsp. cloacae as E. bugandensis or E. ludwigii strains (6,8,9). In contrast, the plasmid-encoded genes (such as blaIMI-2 or blaIMI-6) were mostly identified on a IncFII(Yp)-type plasmid in E. asburiae isolates (3,6,10). We characterized a large collection of IMI/NmcA producers collected in France.
We included all nonduplicate IMI-producing and NmcA-producing isolates showing antimicrobial resistance received at the French National Reference Center for Antimicrobial resistance (F-NRC) during 2012–2022 (n = 112) (Appendix 1 Table 1). Mass spectrometry showed that all strains belonged to the ECC. Since 2014, each year, 3–20 IMI/NmcA producers were identified, representing 0.03%–0.91% of all carbapenemase-producing Enterobacterales analyzed at F-NRC. No IMI/NmcA producers were found before 2014. (Appendix 2 Figure 1).
Disc diffusion antimicrobial susceptibility testing revealed resistance to third-generation cephalosporins for 1 strain (257D9, overexpression of ampC confirmed with CLOXA agar) of the 112 tested. We determined MICs for last-resort antibiotics against highly resistant bacteria on 30 IMI/NmcA producers belonging to several sequence types (STs) (Appendix 1 Table 2). Relebactam restored imipenem activity for 67% of the strains and vaborbactam restored susceptibility to meropenem for all strains with lower MICs than imipenem/relebactam. Then, 37% of the tested strains were susceptible to colistin. All 30 IMI/NmcA producers remained susceptible to cefepime, cefiderocol, and ceftazidime/avibactam.
We performed WGS on the 112 IMI/NmcA producers and identified 74 IMI-1 producers (Appendix 2 Figures 1, 2). Of those, 44 IMI-1–producing ECC were involved in an outbreak in Mayotte and La Réunion islands.
Figure
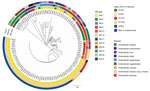
Figure. Phylogenetic relationship and global characterization of 112 IMI-producing E. cloacaecomplex received by the French National Reference Center, France, 2012–2022. The phylogenetic tree was built with a single-nucleotide polymorphism...
We confirmed ECC species identification using average nucleotide identity (ANI) calculation (Appendix 1 Table 3; Appendix 3). E. cloacae subsp. cloacae was the most prevalent species (n = 56 [50.0%]) (Figure). Multilocus sequence typing (MLST) assigned 42 known unique STs for 105 strains. The 7 remaining isolates belonged to new or undetermined STs. Major STs (>4 isolates) were ST820 (n = 45), ST250 (n = 5), ST657 (n = 5), ST1516 (n = 4), and ST1517 (n = 4) (Figure). Of note, 44 of the ST820 strains corresponded to the strain isolated in the Mayotte/La Réunion outbreak; the last IMI-1 E. cloacae subsp. cloacae of ST820, 193I8, was isolated in Paris and was not clonally related to the outbreak strains. That strain exposed >1,200 single-nucleotide polymorphisms (SNPs) corresponding with the other IMI-1 ECC ST820 isolates from Mayotte or La Réunion.
Genes encoding NmcA, IMI-1, IMI-4, IMI-12, and IMI-13 were localized on the chromosome, whereas those coding for IMI-2, IMI-6, IMI-17, IMI-19, IMI-25, IMI-26 and IMI-27 were carried on plasmids. We characterized genetic environments of blaIMI/NmcA genes using Illumina (https://illumina.com) and MinION long-read (Oxford Nanopore, https://nanoporetech.com) sequencing. All chromosome-encoded blaIMI/NmcA genes were located into a EcloIMEX-type genetic element (Appendix 2 Figure 4, panel A), except blaIMI-13, which possessed a distinct genetic environment (Appendix 2 Figure 4, panel B). We detected already-characterized EcloIMEX-type and 6 new variants, named EcloIMEX-11−16 (Appendix 2 Figure 4, panel A). Those EcloIMEX elements were ≈15–≈39.4-kb long, possessed a highly conserved 5′ region, and were inserted between setB and yieP genes. We observed a strong correlation between blaNmcA and EcloIMEX-1. In contrast, we identified blaIMI-1 on 9 different EcloIMEX elements. We saw no correlation between the Enterobacter species and the type of EcloIMEX. The blaIMI-13 gene was inserted in the chromosome between genes encoding a hypothetical protein and an Inovirus-type Gp2 protein. We identified several complete or partially deleted insertion sequences (IS) close to blaIMI-13 (Appendix 2 Figure 4, panel B); however, the mechanism of blaIMI-13 acquisition is unclear.
All blaIMI-6 genes were carried on a IncFII(Yb)-type plasmid (160–200 kb) (Appendix 1 Table 4). Similarly, blaIMI-2 genes were carried on a IncFII(Yp)-type plasmid for 75% (8/12) of the IMI-2 producers. The plasmidic replicase was not identified in the 4 remaining IMI-2 producers. The long-read sequencing performed on strains producing new IMI variants enabled a more precise identification of plasmid type and size (Appendix 1 Table 3). The close genetic environments of the blaIMI genes included several IS that differed according to the blaIMI variants (Appendix 2 Figure 4). Conjugation experiments performed in E. coli J53 used as recipient strain confirmed those plasmids were conjugative except the 1 carrying blaIMI-17.
We built an SNP matrix for the 44 IMI-1 E. cloacae subsp. cloacae ST820 isolates involved in the Mayotte/La Réunion outbreak to confirm their clonality. Those strains were closely related (1–62 SNPs between 2 isolates). We also performed a Bayesian analysis to estimate the date of the most recent ancestor and the evolutionary rate of that population. We estimated the evolutionary rate of the clone to 3.94 × 10−7 substitutions per site and per year (95% highest posterior density [HPD] 2.50–5.33 × 10−7), corresponding to 1.63 SNPs per genome per year (95% HPD 1.04–2.21 SNPs). The common ancestor of the 44 IMI-1–producing E. cloacae subsp. cloacae ST820 isolates has an estimated date of 1994.7 (95% HPD 1990.8–2000.2) (Appendix 2 Figure 5).
Consistent with previous findings (6,9), our collection of IMI producers included uncommon species of ECC, such as E. cloacae subsp. cloacae, a rarely described species; IMI-1, IMI-2 and IMI-6 were the most prevalent variants. We identified no isolates of E. hormaechei, the most prevalent carbapenemase-producing ECC species (11,12).
Genetic environments and plasmid types of IMI-2 producers identified in this study were similar to those previously described (2,3,13); IncFII(Yp)-type plasmids were most common. The close genetic environment of blaIMI-2 observed in our isolates has been reported on a plasmid identified in E. coli (2). The genetic environment of blaIMI-6 was previously reported in an E. cloacae isolate described by Boyd et al. (6). Regarding the chromosome-encoded IMI and NmcA variants (n = 85), we described a variety of EcloIMEX elements (n = 11) including 6 novel elements; that the same EcloIMEX could be identified in different ECC species suggests that XerC/D recombinases enable the mobility of these blaIMI-/NmcA–carrying EcloIMEX structures specifically between ECC species. Finally, the evolution rate of the IMI-1–producing E. cloacae subsp. cloacae ST820 clone (1.63 SNPs/genome/year) is similar to the 0.5–3 SNPs/year for a genome reported for a population of multidrug-resistant ECC in the United Kingdom (14) and the 2.5–3 SNPs/year for a genome identified for ST171 and ST78 carbapenem-resistant ECC (15).
In conclusion, in IMI/NmcA producers in France, we observed a large diversity of ECC species, STs, genetic supports, and genetic environments. Future work should elucidate why E. cloacae subsp. cloacae is highly prevalent among IMI producers; why blaIMI/NmcA-carrying plasmids were almost always found alone in IMI-producing isolates that always do not carry any other resistance genes; and whether EcloIMEX genetic elements are mobilizable. Clinicians should remain aware of potential antimicrobial resistance among ECC species.
Dr. Emeraud is assistant professor at the INSERM. Her main field of research interest includes epidemiology, genetics, and biochemistry of β-lactamases in Gram negatives.
References
- Rasmussen BA, Bush K, Keeney D, Yang Y, Hare R, O’Gara C, et al. Characterization of IMI-1 beta-lactamase, a class A carbapenem-hydrolyzing enzyme from Enterobacter cloacae. Antimicrob Agents Chemother. 1996;40:2080–6. DOIPubMedGoogle Scholar
- Zhang F, Wang X, Xie L, Zheng Q, Guo X, Han L, et al. A novel transposon, Tn6306, mediates the spread of blaIMI in Enterobacteriaceae in hospitals. Int J Infect Dis. 2017;65:22–6. DOIPubMedGoogle Scholar
- Hopkins KL, Findlay J, Doumith M, Mather B, Meunier D, D’Arcy S, et al. IMI-2 carbapenemase in a clinical Klebsiella variicola isolated in the UK. J Antimicrob Chemother. 2017;72:2129–31. DOIPubMedGoogle Scholar
- Naas T, Oueslati S, Bonnin RA, Dabos ML, Zavala A, Dortet L, et al. Beta-lactamase database (BLDB) - structure and function. J Enzyme Inhib Med Chem. 2017;32:917–9. DOIPubMedGoogle Scholar
- Antonelli A, D’Andrea MM, Di Pilato V, Viaggi B, Torricelli F, Rossolini GM. Characterization of a novel putative Xer-dependent integrative mobile element carrying the blaNMC-A carbapenemase gene, inserted into the chromosome of members of the Enterobacter cloacae complex. Antimicrob Agents Chemother. 2015;59:6620–4. DOIPubMedGoogle Scholar
- Boyd DA, Mataseje LF, Davidson R, Delport JA, Fuller J, Hoang L, et al. Enterobacter cloacae complex isolates harboring blaNMC-A or blaIMI-type class A carbapenemase genes on novel chromosomal integrative elements and plasmids. Antimicrob Agents Chemother. 2017;61:e02578–16. DOIPubMedGoogle Scholar
- Koh TH, Rahman NBA, Teo JWP, La MV, Periaswamy B, Chen SL. Putative integrative mobile elements that exploit the Xer recombination machinery carrying blaIMI-type carbapenemase genes in Enterobacter cloacae complex isolates in Singapore. Antimicrob Agents Chemother. 2017;62:e01542–17.PubMedGoogle Scholar
- Miltgen G, Bonnin RA, Avril C, Benoit-Cattin T, Martak D, Leclaire A, et al. Outbreak of IMI-1 carbapenemase-producing colistin-resistant Enterobacter cloacae on the French island of Mayotte (Indian Ocean). Int J Antimicrob Agents. 2018;52:416–20. DOIPubMedGoogle Scholar
- Octavia S, Koh TH, Ng OT, Marimuthu K, Venkatachalam I, Lin RTP, et al. Genomic study of blaIMI-positive Enterobacter cloacae complex in Singapore over a five-year study period. Antimicrob Agents Chemother. 2020;64:e00510–20. DOIPubMedGoogle Scholar
- Rotova V, Papagiannitsis CC, Chudejova K, Medvecky M, Skalova A, Adamkova V, et al. First description of the emergence of Enterobacter asburiae producing IMI-2 carbapenemase in the Czech Republic. J Glob Antimicrob Resist. 2017;11:98–9. DOIPubMedGoogle Scholar
- Peirano G, Matsumura Y, Adams MD, Bradford P, Motyl M, Chen L, et al. Genomic epidemiology of global carbapenemase-producing Enterobacter spp., 2008–2014. Emerg Infect Dis. 2018;24:1010–9. DOIPubMedGoogle Scholar
- Emeraud C, Petit C, Gauthier L, Bonnin RA, Naas T, Dortet L. Emergence of VIM-producing Enterobacter cloacae complex in France between 2015 and 2018. J Antimicrob Chemother. 2022;77:944–51. DOIPubMedGoogle Scholar
- Rojo-Bezares B, Martín C, López M, Torres C, Sáenz Y. First detection of blaIMI-2 gene in a clinical Escherichia coli strain. Antimicrob Agents Chemother. 2012;56:1146–7. DOIPubMedGoogle Scholar
- Moradigaravand D, Reuter S, Martin V, Peacock SJ, Parkhill J. The dissemination of multidrug-resistant Enterobacter cloacae throughout the UK and Ireland. Nat Microbiol. 2016;1:16173. DOIPubMedGoogle Scholar
- Gomez-Simmonds A, Annavajhala MK, Wang Z, Macesic N, Hu Y, Giddins MJ, et al. Genomic and geographic context for the evolution of high-risk carbapenem-resistant Enterobacter cloacae complex clones ST171 and ST78. MBio. 2018;9:e00542–18. DOIPubMedGoogle Scholar
Figures
Cite This ArticleTable of Contents – Volume 30, Number 6—June 2024
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Cécile Emeraud, Service de Bactériologie-Hygiène, Hôpital de Bicêtre, 78 rue du Général Leclerc, 94275 Le Kremlin-Bicêtre CEDEX, France
Top