Volume 32, Number 2—February 2026
Dispatch
Neospora caninum Infection in Marine Mammals Stranding in Northeastern Pacific Ocean Region
Cite This Article
Citation for Media
Abstract
We used immunohistochemistry and PCR to identify Neospora caninum in 6 infected marine mammal species, including 2 pups, that stranded in the northeastern Pacific Ocean. Our findings suggest the expansion of this parasite’s host range to marine mammals, underscoring the effect of terrestrial pathogens that flow from land to sea.
Researchers using immunohistochemistry and PCR-DNA sequencing analyses have confirmed high infection rates of the protozoan parasites Toxoplasma gondii and Sarcocystis neurona, often as mixed infections, in a range of stranded pinniped, cetacean, and mustelid species (1,2). Parasite transmission is closely linked to land-to-sea pathogen flow (3,4). We identified Neospora caninum, a protozoan pathogen known to affect reproductive fitness in livestock, in 6 different species of marine mammals stranded in the northeastern Pacific Ocean region. The emergence of N. caninum illustrates a third terrestrially sourced parasite (also referred to as a pollutagen) flowing from land to sea to infect marine mammals in this region. This parasite is distinct from previously reported N. caninum–like parasites that circulate between pinnipeds in a marine cycle (5).
In cattle, N. caninum is considered among the most efficient pathogens to be transmitted transplacentally (6). This pathogen is a major contributor of reproductive loss in the dairy industry worldwide. Dogs and wild canids, including foxes, wolves, and coyotes, are among the recognized hosts, both definitive and intermediate, for this parasite. Unlike most coccidian parasites that have a limited host range, N. caninum is increasingly detected in a wide array of terrestrial and avian species (7). Although reports have identified antibodies to N. caninum in prior serosurveys of marine mammals in Australia, Japan, and the United States (8–10), those assays were not validated for wildlife. Cross-reactivity with related coccidian parasites that commonly circulate between marine mammals in a marine cycle may have confounded interpretation of the results (11). We report 8 confirmed cases of N. caninum infection in 6 marine mammal species, including 2 pups and a pregnant female.
Figure 1
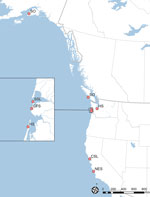
Figure 1. Stranding locations and animal species in study of Neospora caninum infection in marine mammals stranding in the northeastern Pacific Ocean region. NES, northern elephant seal (Mirounga angustirostris...
Throughout the northeastern Pacific region, local and regional marine mammal stranding networks respond to live stranded and dead marine mammals. In this case series, wildlife officials delivered a California sea lion (Zalophus californianus) for rehabilitation. Despite supportive care, the animal declined and was euthanized. We identified a solitary N. caninum infection using PCR, observing no discernible parasites by histopathology. As a result of that finding, we conducted a retrospective database analysis on 410 stranded marine mammals previously screened by PCR-DNA sequencing at the internal transcribed spacer (ITS) 1 marker. We identified 7 additional cases of N. caninum infection in another 5 species of marine mammals. The geographic range of the N. caninum–infected animals (Figure 1) suggested multiple points, rather than a single point source, of parasite exposure.
We documented the stranding location, recorded signalment, compiled morphometrics, and performed a necropsy for all 8 cases (Appendix Table). We also harvested representative tissues for histopathology and immunohistochemistry and froze samples for ancillary diagnostic studies. We conducted PCR-DNA sequencing using pan-Apicomplexan primers that amplify across the ITS1 region for speciation. We carried out immunohistochemistry for N. caninum, T. gondii, and S. neurona on available tissues from 4 PCR-positive animals, following previously reported protocols (1,12).
Figure 2
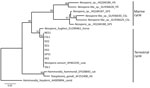
Figure 2. Phylogenetic relationships among Neospora caninum–like species that circulate in a marine cycle compared with terrestrial-sourced species from study of N. caninuminfection in marine mammals stranding in...
Phylogenetic analysis at the ITS1 locus established unequivocally that all 8 animals were infected with an identical sequence type that resolved as N. caninum (GenBank accession nos. PX529932–8) (Figure 2). For comparison, we included in the tree sequences recovered from other pinnipeds infected by N. caninum–like parasites that commonly circulate within a marine transmission cycle (5). We conducted PCR testing, determining 7 of the 8 N. caninum–infected animals had polyparasite infections with 1 or 2 other terrestrially sourced coccidian agents (T. gondii, S. neurona) in their tissues. Infected animals were 2 sea otters (Enhydra lutris), 2 harbor seals (Phoca vitulina), 1 northern elephant seal (Mirounga angustirostris), 1 Guadelupe fur seal (Arctocephalus townsendi), and 1 Steller sea lion (Eumetopias jubatus) (Appendix Table).
Figure 3
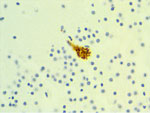
Figure 3. Immunohistochemistry of a harbor seal pup with Neospora caninum antigen localized within a neuron from study of N. caninuminfection in marine mammals stranding in the northeastern...
We diagnosed nonsuppurative and necrotizing encephalitis (n = 2), meningoencephalomyelitis (n = 1), and meningoencephalitis (n = 1) by histology of the brain for 4 of 6 cases, including the pup and subadult harbor seals, adult Steller sea lion, and yearling California sea lion. We conducted immunohistochemistry in 4 encephalitic cases, observing N. caninum antigen in brain samples from the harbor seal pup and T. gondii and S. neurona (one or both) antigens in the other 3 animals (Figure 3). We found no pathognomonic lesions in this case series directly attributed to N. caninum infection. The cause of death for all 8 animals was independent of N. caninum infection (Appendix Table).
We identified 8 animals from 6 marine mammal species unequivocally infected with N. caninum. Infection by this parasite alone was confirmed in 1 animal, a yearling California sea lion, and the infection represents another example of land-to-sea flow of a pollutagen of parasite origin. The contribution of this pathogen to marine mammal health is unknown. All infected animals we studied were from species with varying degrees of philopatry, with either short natal dispersal distances or extended oceanic to pelagic migrations. Seasonal haul out associated with pupping and breeding, coupled with accounts of wild canids scavenging at rookeries (13), provide insights into potential parasite introduction, persistence, and dissemination. For sea otters, bioaccumulation of parasites by invertebrate prey species may be inferred from observations of transmission dynamics with T. gondii (4). On the basis of gross and microscopic findings, we determined N. caninum infection to be unrelated to the proximate cause of death in the examined animals. Nevertheless, detection of N. caninum in a pregnant Steller sea lion, as well as in a harbor seal pup and northern elephant seal pup (weanling), is concerning. Recrudescence of latent infection during pregnancy contributes to parasite reactivation that results in vertical transmission in cattle and dogs (6), and we propose this phenomenon might be occurring sporadically in marine mammals.
Documented reports have confirmed land-to-sea transmission of protozoan parasites, and serosurveys for N. caninum have detected titers in marine mammal species. A recent review documents the global extent of land-to-sea pathogen flow (14). Seals and sea lions are monophyletic, members of the order Carnivora, and share a common ancestry with terrestrial canids. This evolutionary relationship may predispose these marine mammals to pollutagens defecated by canids. Cross reactivity with previously described nonpathogenic N. caninum–like parasites that circulate among pinnipeds in a marine transmission cycle, including coccidia types A and B (with California sea lions as definitive hosts) (5), may have contributed to false-positive results in prior serosurveys. The contribution of these N. caninum–like parasites to immune protection of hosts against N. caninum is unknown. In this limited series, we confirmed N. caninum infection in 2 sea otters, 2 harbor seals, 1 California sea lion, 1 northern elephant seal, 1 Steller sea lion, and 1 Guadelupe fur seal, none of which had been infected with previously described N. caninum–like parasites. Those findings extend the host range and ecologic niche for N. caninum. Infections were predominantly mixed with T. gondii, S. neurona, or both. Climate change, ecologic marine regime shifts, rural and urban development, weather events, and other environmental perturbations may lead to incursion of previously recognized land-based pathogens into the marine environment.
In conclusion, our results indicate that further investigations to characterize the life history of N. caninum in the marine environment, the role of polyparasitism in disease manifestation, and the potential pathogenicity in susceptible host species are warranted. Whereas we cannot definitively attribute N. caninum as the cause of illness or death for the animals described here, future investigations of similar unexpected deaths, particularly those involving abortion storms among coastal breeding marine mammals in the northeastern Pacific, should consider molecular screening for this pathogen.
Dr. Raverty is a veterinary pathologist at the Animal Health Center, British Columbia Ministry of Agriculture, Abbotsford, British Columbia, Canada. He has investigated infectious and noninfectious disease entities in live and dead stranded marine mammals throughout the northeastern Pacific.
Acknowledgments
We gratefully acknowledge the postmortem room and histology staff at the Animal Health Center.
We obtained Letters of Authorization from Regional Stranding Coordinators, Convention on International Trade in Endangered Species permits, National Oceanic and Atmospheric Administration (TMMC permit no. 26532), and United States Fish and Wildlife Service declarations for this investigation.
Prescott Awards to P.D. and D.M.L. funded this project. This research was supported in part by the Intramural Research Program of the National Institutes of Health (NIH). The contributions of the NIH author(s) were made as part of their official duties as NIH federal employees, are in compliance with agency policy requirements, and are considered Works of the United States Government. However, the findings and conclusions presented in this paper are those of the author(s) and do not necessarily reflect the views of the NIH, the US Department of Health and Human Services, or any other Agency of the US Government.
References
- Gibson AK, Raverty S, Lambourn DM, Huggins J, Magargal SL, Grigg ME. Polyparasitism is associated with increased disease severity in Toxoplasma gondii-infected marine sentinel species. PLoS Negl Trop Dis. 2011;5:
e1142 . DOIPubMedGoogle Scholar - Miller MA, Shapiro K, Murray MJ, Haulena M, Raverty SA. Protozoan parasites of marine mammals. In: Gulland FMD, Dierauf LA, Whitman KL, editors. CRC handbook of marine mammal medicine, 3rd edition. Oxford: CRC Press; 2018;chap 20:p. 1-46.
- Burgess TL, Tinker MT, Miller MA, Smith WA, Bodkin JL, Murray MJ, et al. Spatial epidemiological patterns suggest mechanisms of land-sea transmission for Sarcocystis neurona in a coastal marine mammal. Sci Rep. 2020;10:3683. DOIPubMedGoogle Scholar
- Conrad PA, Miller MA, Kreuder C, James ER, Mazet J, Dabritz H, et al. Transmission of Toxoplasma: clues from the study of sea otters as sentinels of Toxoplasma gondii flow into the marine environment. Int J Parasitol. 2005;35:1155–68. DOIPubMedGoogle Scholar
- Carlson-Bremer D, Johnson CK, Miller RH, Gulland FM, Conrad PA, Wasmuth JD, et al. Identification of two novel coccidian species shed by California sea lions (Zalophus californianus). J Parasitol. 2012;98:347–54. DOIPubMedGoogle Scholar
- Dubey JP, Schares G, Ortega-Mora LM. Epidemiology and control of neosporosis and Neospora caninum. Clin Microbiol Rev. 2007;20:323–67. DOIPubMedGoogle Scholar
- Dubey JP, Zarnke R, Thomas NJ, Wong SK, Van Bonn W, Briggs M, et al. Toxoplasma gondii, Neospora caninum, Sarcocystis neurona, and Sarcocystis canis-like infections in marine mammals. Vet Parasitol. 2003;116:275–96. DOIPubMedGoogle Scholar
- Omata Y, Umeshita Y, Watarai M, Tachibana M, Sasaki M, Murata K, et al. Investigation for presence of Neospora caninum, Toxoplasma gondii and Brucella-species infection in killer whales (Orcinus orca) mass-stranded on the coast of Shiretoko, Hokkaido, Japan. J Vet Med Sci. 2006;68:523–6. DOIPubMedGoogle Scholar
- Wong A, Lanyon JM, O’Handley R, Linedale R, Woolford L, Long T, et al. Serum antibodies against Toxoplasma gondii and Neospora caninum in southeast Queensland dugongs. Mar Mamm Sci. 2020;36:180–94. DOIGoogle Scholar
- Donahoe SL, Lindsay SA, Krockenberger M, Phalen D, Šlapeta J. A review of neosporosis and pathologic findings of Neospora caninum infection in wildlife. Int J Parasitol Parasites Wildl. 2015;4:216–38. DOIPubMedGoogle Scholar
- Barbosa L, Johnson CK, Lambourn DM, Gibson AK, Haman KH, Huggins JL, et al. A novel Sarcocystis neurona genotype XIII is associated with severe encephalitis in an unexpectedly broad range of marine mammals from the northeastern Pacific Ocean. Int J Parasitol. 2015;45:595–603. DOIPubMedGoogle Scholar
- Reid RE, Gifford-Gonzalez D, Koch PL. Coyote (Canis latrans) use of marine resources in coastal California: a new behavior relative to their recent ancestors. Holocene. 2018;28:1781–90. DOIGoogle Scholar
- Song L, Hu C, Zhang X, Liu Y, Song G, Wu J, et al. Effects of feeding on microbial community structure and pathogen abundance in marine aquaculture ponds. Mar Environ Res. 2025;210:
107319 . DOIPubMedGoogle Scholar
Figures
Cite This ArticleOriginal Publication Date: February 17, 2026
1Current affiliation: National Oceanic and Atmospheric Administration Fisheries, PRD/Alaska Region, US Department of Commerce, Anchorage, Alaska, USA.
Table of Contents – Volume 32, Number 2—February 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Michael E. Grigg, Laboratory of Parasitic Diseases, NIAID National Institutes of Health, 4 Center Dr, Rm B1-06, Bethesda, MD 20892 USA
Top