Volume 8, Number 3—March 2002
Research
Molecular Epidemiology of Adenovirus Type 7 in the United States, 1966–20001
Cite This Article
Citation for Media
Abstract
Genetic variation among 166 isolates of human adenovirus 7 (Ad7) obtained from 1966 to 2000 from the United States and Eastern Ontario, Canada, was determined by genome restriction analysis. Most (65%) isolates were identified as Ad7b. Two genome types previously undocumented in North America were also identified: Ad7d2 (28%), which first appeared in 1993 and was later identified throughout the Midwest and Northeast of the United States and in Canada; and Ad7h (2%), which was identified only in the U.S. Southwest in 1998 and 2000. Since 1996, Ad7d2 has been responsible for several civilian outbreaks of Ad7 disease and was the primary cause of a large outbreak of respiratory illness at a military recruit training center. The appearance of Ad7d2 and Ad7h in North America represents recent introduction of these viruses from previously geographically restricted areas and may herald a shift in predominant genome type circulating in the United States.
Human adenoviruses (Ads) comprise 51 serotypes (1); they are ubiquitous and responsible for a wide range of clinical syndromes. Among recognized serotypes, Ad type 7 (Ad7) (and to a lesser extent Ad type 3) is most often associated with severe disease (2). Although Ad7 infections typically result in mild upper respiratory tract illnesses and conjunctivitis, infections can also lead to more serious lower respiratory tract illnesses, disseminated disease, and death, particularly in infants and persons with underlying immunologic or respiratory compromise (3–7). Ad7 infections have also been associated with diseases of the central nervous system (8,9) and long-term respiratory sequelae that include bronchiectasis and hyperlucent lung or McLeod syndrome (10).
Ad7 accounts for nearly 20% of all Ads reported to the World Health Organization (11), and family clusters and institutional and communitywide outbreaks of Ad7 disease have been extensively documented (5,12–18). Three types of outbreaks have been described (12): i) outbreaks that occur during the winter months among institutionalized infants (<2 years of age) that result in high rates of severe illness and death; ii) periodic nonseasonal communitywide outbreaks involving older children and adults with infrequent serious outcomes; and iii) outbreaks of acute respiratory disease among new military recruits. Outbreaks of acute respiratory disease due primarily to Ad7 and Ad4 were an important cause of illness in new military recruits in the United States until live enteric-coated Ad4 and Ad7 vaccines began to be routinely administered in 1971 (19). The recent cessation of production and administration of these vaccines has resulted in a resumption of Ad-associated acute respiratory disease outbreaks at military recruit training centers throughout the United States (20–22).
To facilitate study of the molecular epidemiology of Ad7, a classification system based on restriction enzyme analysis of Ad genomic DNA was devised by Li and Wadell (23) and later revised by Li et al. (24). Their system uses BamHI as the "type" defining enzyme, with different genome types denoted with a character, e.g., “p” for the Ad7 prototype strain, Gomen; and then “a” through “k.” Genome types that are further distinguished by restriction pattern with additional selected enzymes are given an Arabic numeral (e.g., Ad7p, p1, a, a1-6). Their system has been widely used to correlate genome types with geographic distribution and pathogenic potential.
Both globally dispersed and geographically restricted genome types of Ad7 have been identified by restriction analysis, and regional shifts or replacements of predominant genome types have been documented on different continents. Among the 3 Ad7 genome types first distinguished by restriction analysis (25), two shown to be serologically distinct (26) were designated Ad7p (Gomen) and Ad7a (S-1058), and a third, designated Ad7b, was thought to be associated with more severe illness (12). Ad7b eventually spread worldwide (27–30), displacing formerly common genome types (i.e., Ad7p, Ad7a, Ad7a1-6, Ad7c, and others) that are now rarely detected. Exceptions to this pattern have been reported. In the former Soviet Union, a successive shift from Ad7a and Ad7a1-5 to Ad7f1 during 1976-1979 and 1986-1988 was reported (31). In South America, a shift from Ad7c to Ad7h occurred in 1986 (32), and Ad7h has subsequently caused serious respiratory illness in infants and young children in Chile and Argentina (33,34). In the early 1980s in China, a new genome type, Ad7d (27), replaced Ad7b as the predominant circulating virus. Recent reports suggest that Ad7d and Ad7h have spread beyond their formerly geographically restricted regions. Ad7d was identified in Japan in 1987 (35), and countrywide epidemics of Ad7 that began in 1995 in Korea (18; Hoan-Jong Lee, pers. comm.) and Japan (15,36) were attributed to Ad7d and a closely related genomic variant, Ad7d2, respectively. Ad7d2 has emerged as the predominant strain circulating in Israel since 1992 (37). Ad7h was first reported outside South America in 1996, in Japan (36,38).
Beginning in the fall of 1998, an outbreak of Ad7 infection occurred at a pediatric chronic-care facility in Chicago and subsequently spread to a tertiary-care hospital, where staff from two clinic units were infected (17). This multi-site outbreak was associated with considerable illness and death among residents of the chronic-care facility. Isolates from this outbreak were identified by restriction enzyme analysis as Ad7d2. The appearance of this new genome type prompted us to study the temporal and geographic distribution of Ad7 genome types in the United States to better characterize the emergence and spread of this virus.
Ads
Of 297 Ad field isolates obtained from the Centers for Disease Control and Prevention (CDC) archives, state public health laboratories, university hospitals, and military training centers, 166 confirmed as Ad7 were selected for genome type analysis (Table 1). Of these, 116 were obtained from 1966 to 2000 from civilians in 25 states and eastern Ontario, Canada; 50 were obtained from February 1997 to May 1998 from military recruits attending training centers in five states (20). Isolates were selected to achieve broad geographic and temporal distribution. Because detailed demographic, epidemiologic, and clinical data from patients were limited, they were not included in this report. Most civilian isolates were obtained from individual cases or family clusters of Ad7 disease, ranging from mild upper respiratory illness to severe lower respiratory tract illness and death. Where civilian outbreaks of Ad7 illness were recognized, only one representative isolate was included in the 166 sample for analysis. Approximately 10% of Ad7 isolates from military recruits with respiratory illness were sampled from all five training sites and were selected to be evenly spaced over the designated time period. Reference strains Gomen and S-1058 were obtained from the American Type Culture Collection (ATCC, Rockville, MD). Reference isolates of Ad7d2 were obtained from a postmortem rectal swab from a 4-month-old baby in Israel in 1993, and Ad7h was obtained during a regional outbreak of respiratory illness in Chile in 1998. All Ad isolates were passaged at least once in A549 cells before restriction analysis. Type-specificity of all Ad7 field isolates was confirmed by neutralization or Ad7 type-specific polymerase chain reaction assay (PCR) (39).
DNA Restriction Analysis
Ad genomic DNA was extracted by a modification of the method of Deryckere and Burgert (40). Briefly, isolates were grown in 75-cm2 confluent flasks of A549 cells until the 4+ stage of cytopathic effect was attained. The contents of the flask were centrifuged at low speed to remove cells, and the supernatant was transferred to an ultra-centrifuge tube and centrifuged for 2 hours at 100,000 x g. The virus pellet was resuspended in 400 μL of Tris buffer (pH 7.4) with 1% sodium dodecyl sulfate and sequentially digested with DNAse free RNAse A (0.1 mg/mL) and proteinase K (0.5 mg/mL). The digest was extracted twice with equal volumes of phenol and chloroform/isoamylalcohol (24:1) and once with chloroform/isoamylalcohol alone. The purified DNA was then precipitated with absolute ethanol and washed once with 75% ethanol, and the pellet was resuspended in 100 μL of dH2O. Enzyme digestions were carried out according to manufacturer's instructions (Boehringer Mannheim Biochemicals, Indianapolis, IN). DNA from all Ad7 isolates was digested with BamHI and SmaI, and selected isolates were also digested with enzymes BcII, BglI, BglII, BstEII, EcoRI, HpaI, HindIII,SalI, XbaI, and XhoI. Enzyme digests were electrophoresed at 100 volts for 5 hours on 0.8% agarose gels, and the DNA bands were visualized by ethidium bromide staining. Restriction fragment size(s) was interpolated from DNA molecular weight standards included in each run. Restriction patterns were compared with previously published profiles (24,37,41,42), and the identification of genome types followed the denomination system of Li et al. (24).
DNA Sequencing
The hypervariable region of the hexon protein gene corresponding to nucleotides 403 to 1356 (Gomen), which have been shown to encode the residues that define Ad serotype, was PCR amplified from selected Ad7 isolates as described (43) and sequenced by using the DyeDeoxy Terminator Cycle Sequencing Kit and ABI 373A automated DNA sequencer (Applied Biosystems, Foster City, CA). Nucleotide sequences were determined for both PCR product strands. Sequence analysis was performed by using the Wisconsin Package ver. 10.0 (Genetics Computer Group, Madison, WI). Hexon gene sequence data for the reference Ad7d2 strain from Israel were submitted to GenBank (accession number AF321311).
Ad7 Genome Types Identified
Figure
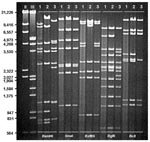
Figure. Restriction profiles of representative human adenovirus (Ad) genome types Ad7b (1), Ad7d2 (2), and Ad7h (3) after digestion with selected enzymes, BamHI, Sma I, BstEII, BglII, and BcII. DNA markers II...
DNA restriction analysis of the 166 Ad7 field isolates identified 108 (65%) as Ad7b, 46 (28%) as Ad7d2, 4 (2%) as Ad7h, 3 (2%) as Ad7p, 3 (2%) as Ad7a, and 2 (1%) as Ad7a3. Restriction profiles of representative Ad7b, Ad7d2, and Ad7h isolates for selected endonucleases are shown in the Figure. All U.S. (and eastern Ontario, Canada) Ad7d2 isolates and an Ad7d2 reference strain from Israel (37) gave identical restriction patterns for BamHI, BcII, BglI, BglII, BstEII, EcoRI, HpaI, HindIII, SalI, SmaI, XbaI, and XhoI. Identical restriction profiles were also obtained with four U.S. Ad7h isolates and a 1998 isolate of Ad7h from Chile, which were similar to profiles described for Ad7h strains isolated in Argentina and Chile (formerly designated Ad3f) (41,42,44).
Ad7 Hexon Gene Sequencing
The hypervariable regions of the hexon gene (corresponding to nucleotides 403 to 1356 of the reference strain Gomen) of 24 temporally and geographically diverse Ad7 field isolates (including 11 Ad7b, 10 Ad7d2, and 3 Ad7h and laboratory strains S-1058, 55142 vaccine, and Gomen) were sequenced and compared with published Ad7 hexon sequences available from GenBank (Table 2). Nucleotide and deduced amino acid alignments of these sequences comprised two major genetic clusters as previously described (26,45): cluster 1, Ad7p (Gomen) and Ad7p1; and cluster 2, Ad7a Ad7b, Ad7c, Ad7d, Ad7d2, Ad7g, and Ad7h. Cluster 2 sequences were highly conserved, with over 98% nucleotide identity, and were generally uncorrelated with genome type. However, a unique Gln substitution for Leu (codon CTG > CAG) at amino acid position 443 of loop 2 of the predicted hexon protein was identified in all 10 Ad7d2 isolates from the United States and Israel; this substitution was also present in published hexon sequences of Ad7d isolates from China (45) and Japan (38).
Temporal Distribution of Ad7 Genome Types
The yearly distribution of the 166 Ad7 genome types is shown in Table 3. Ad7b was the only genome type identified from 1970 through 1992 and was the predominant genome type identified through 2000. Ad7d2 first appeared among 1993 isolates and accounted for approximately 28% of all Ad7 isolates obtained from 1993 to 2000. Four epidemiologically unrelated isolates of Ad7h were identified in 1998 and 2000.
Geographic Distribution of Ad7 Genome Types
Ad7b was identified among isolates from nearly all states (and eastern Ontario) sampled. Ad7d2 was first identified in isolates from Maryland and New York in 1993 and thereafter primarily from midwestern and northeastern states, including Wisconsin, Illinois, Kansas, Missouri, Louisiana, South Carolina, and Ohio, as well as eastern Ontario. Ad7h was only identified among isolates obtained from Texas and Arizona.
Ad7 Civilian Outbreaks, 1996-2000
During this study, we became aware of five separate outbreaks of Ad7 respiratory illness among civilians (Table 4). Four were institutional outbreaks that involved primarily infants and young children with underlying chronic disease that occurred in the fall or summer months of 1996, 1998, 1999, and 2000. A fifth communitywide outbreak of Ad7 in Tennessee, which occurred during March-July 1997, involved previously healthy children (16).Genome type analysis at CDC identified Ad7d2 in three of the four outbreaks where isolates were available. We attributed one outbreak to a novel Sma I restriction variant of Ad7b (Ad7bvar) that occurred in New York in 1999 (Jennifer Calder, manuscript in preparation).
Ad7 Gnome Types at Military Recruit Training Centers
In anticipation of increased Ad activity following termination of routine vaccination of new military recruits in 1996, the Naval Health Research Center (NHRC) in San Diego, CA, initiated surveillance for new cases of Ad-associated respiratory illness (20) [http://www.nhrc.navy.mil/geis/sites/nhrc.htm]. Clinical specimens collected at five designated training centers (San Diego, CA; San Antonio, TX; St. Robert, MO; Great Lakes, IL; and Columbia, SC) from recruits who reported respiratory illness were submitted to NHRC for identification of viral and bacterial pathogens. Of 50 Ad7 isolates, Ad7d2 was the most common genome type identified (58%), followed by Ad7b (34%), Ad7p (6%), and Ad7h (2%) (Table 5). Most Ad7d2 infections were reported from the Naval Recruit Training Center in Great Lakes, IL, where an outbreak of Ad7 (and Ad3) respiratory illness was documented during the fall of 1997 (21). Over 70% of the Ad7 isolates sampled from the Great Lakes Center from September 1997 to February 1998 were identified as Ad7d2. One of four Ad7h isolates identified in this study was obtained from a new recruit at Lackland Air Force Base, in San Antonio.
Our study represents the most comprehensive survey to date of Ad7 genome types circulating in the United States and provides a basis for future surveillance studies that can better delineate the disease impact of these viruses.
Before this study, the most comprehensive surveys of Ad7 genomic variants in the United States were conducted by Wadell et al. (27) and Adrian et al. (28) with field isolates of Ad7 collected from 1961 to 1985. These authors identified a diverse group of cocirculating Ad7 genome types (Ad7p, Ad7a, Ad7c, and others) that by the late 1960s to early 1970s were replaced by Ad7b, a change that preceded similar shifts to Ad7b seen in other parts of the world. Our data confirm this observation and show a continued dominance of the Ad7b genome type in the United States. Only one genome type from the earlier period, Ad7p, was still identified among currently circulating strains. We also documented the appearance of two new Ad7 genome types: Ad7d2, which was first identified in specimens collected in 1993 and subsequently detected over a wide geographic area in the eastern half of the United States and Canada; and Ad7h, which was first identified in specimens collected in 1998 in the Southwest.
Both epidemiologic and molecular evidence suggests that Ad7d2 entered the United States as part of its recent spread from evolutionarily related Ad7d strains formerly restricted to China. Ad7d2 shows the highest degree of genetic relatedness to Ad7d, differing by only one BstEII restriction site in pairwise comigrating restriction fragment analysis with 12 different endonucleases (24,37); it possesses the unique amino acid substitution in the hexon protein also present in Ad7d isolates from China (45) and Japan (38). Ad7d was identified as early as 1980 in Beijing (24) and 2 years later in Changchin (46), and rapidly displaced Ad7b to become the major genome type circulating in China through 1990. Ad7d was identified in Japan during 1987 to 1992 (35) and in Korea in 1995 (18; Hoan-Jong Lee, pers. comm.), and Ad7d2 was the predominant genome type isolated during the 1995-1998 Ad7 epidemic in Japan (15,36). Ad7d2 was subsequently identified in Israel in 1992 (37) and in the United States in this study in 1993.
The emergence and apparent global spread of Ad7d2 are reminiscent of observations for another genome type of serotype 7, Ad7b. Originally described by Wadell and Varsanyi (25), Ad7b was associated with outbreaks of severe respiratory illness in Europe in the 1970s (12). Although first isolated in 1956 from a Paris orphanage outbreak (12,47), subsequent retrospective studies did not identify Ad7b in Europe again until 1969 (27). Before then, the earliest documented occurrence of Ad7b was in China in 1958 (24), where it was the predominant genome type circulating through the early 1980s (24,46). With the exception of Paris, the first appearance of Ad7b outside China was on the U.S. West Coast in 1962 (27). By 1970, Ad7b was the predominant genome type circulating throughout the United States (28) and eventually throughout many parts of the world.
The mechanism(s) underlying the apparent greater fitness of some Ad7 genome types, as reflected by their capacity to displace other circulating strains, remains speculative. Possible explanations include mutations or recombinations that yield strains with increased pathogenicity and therefore greater chance of causing recognized illness, or biological or antigenic changes that enhance transmission or infection compared with other Ad7 genome types. Although there is no conclusive evidence of differences in pathogenicity between Ad7 genome types, some types appear to be more frequently isolated from healthy carriers (e.g., Ad7p and Ad7a), while others are more often isolated from patients with more serious clinical outcomes (e.g., Ad7b, Ad7c, Ad7d, and Ad7h) (27,34). Some antigenic differences between Ad7 genome types have also been demonstrated; recent studies identified minor differences in neutralization titer between Ad7 prototype strain Gomen (Ad7p) and the vaccine strain 55142 (Ad7a) with rabbit hyperimmune antisera (26). In addition, a unique amino acid substitution in the hexon protein that distinguishes Ad7d/Ad7d2 strains from other genome types is predicted to impart substantial changes in the hydrophilicity of the protein and possibly associated antigenic changes (45).
Although Ad7 can be spread directly by the respiratory route, efficiency of transmission is typically lower than for some other respiratory viruses. Efficient spread usually requires crowding, such as that in closed communities like chronic-care facilities, military barracks, and day-care centers. Widespread community outbreaks of Ad7 can occur but appear to require low levels of herd immunity. For example, in Japan, >95% of persons <40 years of age lacked specific antibodies to Ad7 before the countrywide epidemic of Ad7 that began in 1995 (48,49). The most comprehensive recent seroprevalence data on Ad7 in the United States were obtained in 1992 from 364 military basic trainees attending new recruit training centers (50). Approximately 73% of screened trainees lacked specific antibodies to Ad7. In another study to evaluate the potential for use of Ad vectors in gene therapy for cystic fibrosis, 73.9% of 46 serum specimens collected from 1993 to 1995 from children (median age 4.7 years) were seronegative for Ad7 (51).
To achieve rapid spread, a novel genome type presumably requires an immunologically naive population, greater biological fitness than the indigenous circulating strains, and a means of introduction to the susceptible community. Azar et al. (37) noted that the appearance of Ad7d2 in Israel coincided with the arrival of large numbers of immigrants from the former Soviet Union and Ethiopia during the early 1990s. The global spread of Ad7b in the 1960s and 1970s may have been aided by the movement of unvaccinated U.S. and allied military personnel during the Vietnam War. In our study, the appearance of Ad7h in the U.S. Southwest in 1998 may be explained by the emigration of persons from Ad7h-endemic regions of South America, where communitywide outbreaks of respiratory illness due to Ad7h occurred as recently as 1998 in Chile (Rodrigo Fasce, pers. comm.). However, a more comprehensive survey of Ad7 isolates from Mexico and U.S. states on the Mexican Border would be necessary to substantiate this observation.
The five recognized civilian outbreaks of Ad7 respiratory illness that occurred during 1996-2000, three of which we attributed to genome type 7d2, might have been due to increased reporting as a result of our interest in this study or may represent a real increase in Ad7-associated disease, as occurred in Europe during the early 1970s and in Japan and Korea (12,15,18) beginning in 1995. One unsubstantiated possibility is that the discontinuation of vaccination of U.S. military recruits for Ad4 and Ad7 in 1996 and the subsequent increase in Ad-associated disease at military bases throughout the United States (20–22) provided a new focus for Ad7 dissemination to civilian populations. A possible example of this is the 1998 outbreak of Ad7d2 illness at a Chicago pediatric chronic-care facility described earlier (17). This outbreak occurred within a few miles of the Naval Training Center in Great Lakes, which had had an outbreak of Ad7d2 the preceding year (21). Most cases of Ad infection at military bases since 1996 have been attributed to Ad4 (20,22), but no comparable outbreaks of Ad4 disease among civilians have been reported. Unlike Ad7, which poses a risk to both civilian and military populations, Ad4 has only infrequently been associated with outbreaks of respiratory illness in civilian populations (2).
Although we identified individual cases of severe lower respiratory tract illness and deaths attributed to Ad7d2 and Ad7h in this study, the possibility that these two genome types may be associated with more severe disease is not yet clear. More extensive clinical and epidemiologic study is required to adequately address this question. The limited data from infected military recruits suggest no differences in clinical illness between those infected with Ad7d2 and Ad7b (data not shown). Reports of Ad7d2 infections in Israel (37) and Ad7d infections in China (46) also noted no clear differences in severity of disease. Cases of severe pneumonia and neurologic disease were reported from a recent regional epidemic of Ad7d2 in Japan (15), but there was no evidence that these severe cases were more common than those reported for outbreaks involving other Ad7 genome types. Ad7h, a genetically unique recombinant between Ad7 and Ad3 (42), has been linked to increased illness and death in infants in Chile and Argentina, where it is second only to Human respiratory syncytial virus as a cause of severe viral pneumonia in infants and young children (34,52). However, in this study, too few cases of Ad7h infection were identified to assess differences in disease severity.
In conclusion, our study documents the recent appearance in the United States of two new Ad7 genome types, Ad7d2 and Ad7h, and provides additional evidence of the global spread of these formerly geographically restricted viruses. The possibility that these genome types may be associated with more severe disease makes it prudent to monitor their spread and associated disease.
Dr. Erdman is acting chief of the Respiratory Virus Section in the Division of Viral and Rickettsial Diseases, Centers for Disease Control and Prevention. His interests include development of diagnostic assays and molecular epidemiology of respiratory viruses.
Acknowledgment
We thank those who kindly provided human adenovirus isolates for this study: Richard Buller, Nando Chatterjee, Ronald Cheshier, Pam Colarusso, Rodrigo Fasce, Steve LaCroix, Jane LaFlash, Gary Leonardi, Marrietta Malasig, Ella Mendelson, Roswitha Milk, Laura Montague, Mahin Park, MaryAnn Patterson, Iqbal Poshni, Bill Reimels, Syed Sherazi, Suzanne Spencer, Marsha Tolson, and Cindi Ventrola.
References
- De Jong JC, Wermenbol AG, Verweij-Uijterwaal MW, Slaterus KW, Wertheim-Van Dillen P, Van Doornum GJ, Adenoviruses from human immunodeficiency virus-infected individuals, including two strains that represent new candidate serotypes Ad50 and Ad51 of species B1 and D, respectively. J Clin Microbiol. 1999;37:3940–5.PubMedGoogle Scholar
- Hierholzer J. Adenoviruses. In: Lennette EH, Lennette DA, Lennette ET, editors. Diagnostic procedures for viral, rickettsial and chlamydial infections. 7th ed. Washington: American Public Health Association; 1995. p. 169-88.
- Simila S, Ylikorkala O, Wasz-Hockert O. Type 7 adenovirus pneumonia. J Pediatr. 1971;79:605–11. DOIPubMedGoogle Scholar
- Field PR, Patwardhan J, McKenzie JA, Murphy AM. Fatal adenovirus type 7 pneumonia in an adult. Med J Aust. 1978;2:445–7.PubMedGoogle Scholar
- Alpert G, Charney E, Fee M, Plotkin SA. Outbreak of fatal adenoviral type 7a respiratory disease in a children's long-term care inpatient facility. Am J Infect Control. 1986;14:188–90. DOIPubMedGoogle Scholar
- Zarraga AL, Kerns FT, Kitchen LW. Adenovirus pneumonia with severe sequelae in an immunocompetent adult. Clin Infect Dis. 1992;15:712–3.PubMedGoogle Scholar
- Munoz FM, Piedra PA, Demmler GJ. Disseminated adenovirus disease in immuno-compromised and immunocompetent children. Clin Infect Dis. 1998;27:1194–1200. DOIPubMedGoogle Scholar
- Ladisch S, Lovejoy FH, Hierholzer JC, Oxman MN, Strieder D, Vawter GF, Extrapulmonary manifestations of adenovirus type 7 pneumonia simulating Reye syndrome and the possible role of an adenovirus toxin. J Pediatr. 1979;95:348–55. DOIPubMedGoogle Scholar
- Sly PD, Soto-Quiros ME, Landau LI, Hudson I, Newton-John H. Factors predisposing to abnormal pulmonary function after adenovirus type 7 pneumonia. Arch Dis Child. 1984;59:935–9. DOIPubMedGoogle Scholar
- Schmitz H, Wigand R, Heinreich W. Worldwide epidemiology of human adenovirus infections. Am J Epidemiol. 1983;117:455–66.PubMedGoogle Scholar
- Wadell G, Varsanyi TM, Lord A, Sutton RN. Epidemic outbreaks of adenovirus 7 with special reference to the pathogenicity of adenovirus genome type 7b. Am J Epidemiol. 1980;112:619–28.PubMedGoogle Scholar
- Straube RC, Thompson MA, Van Dyke RB, Wadell G, Connor JD, Wingard D, Adenovirus type 7b in a children's hospital. J Infect Dis. 1983;147:814–9.PubMedGoogle Scholar
- Finn A, Anday E, Talbot GH. An epidemic of adenovirus 7a infection in a neonatal nursery: course, morbidity, and management. Infect Control Hosp Epidemiol. 1988;9:398–404.PubMedGoogle Scholar
- Yamadera S, Yamashita K, Akatsuka M, Kato N, Inouye S. Trend of adenovirus type 7 infection, an emerging disease in Japan. A report of the National Epidemiological Surveillance of Infectious Agents in Japan. Jpn J Med Sci Biol. 1998;51:43–51.PubMedGoogle Scholar
- Mitchell LS, Taylor B, Reimels W, Barrett FF, Devincenzo JP. Adenovirus 7a: a community-acquired outbreak in a children's hospital. Pediatr Infect Dis J. 2000;19:996–1000. DOIPubMedGoogle Scholar
- Gerber SI, Erdman DD, Pur SL, Diaz PS, Segreti J, Kajon A, Outbreak of adenovirus genome type 7d2 infection in a pediatric chronic-care facility and tertiary-care hospital. Clin Infect Dis. 2001;32:694–700. DOIPubMedGoogle Scholar
- Hong J-Y, Lee H-J, Piedra PA, Choi E-H, Park K-H, Koh Y-Y, Lower respiratory tract infections due to adenovirus in hospitalized Korean children: epidemiology, clinical features, and prognosis. Clin Infect Dis. 2001;32:1423–9. DOIPubMedGoogle Scholar
- Top FH Jr. Control of adenovirus acute respiratory disease in U.S. Army trainees. Yale J Biol Med. 1975;48:185–95.PubMedGoogle Scholar
- Gray GC, Goswami PR, Malasig M, Hawksworth AW, Trump DH, Ryan MA, Adult adenovirus infections: loss of orphaned vaccines precipitates military respiratory disease epidemics. Clin Infect Dis. 2000;31:663–70. DOIPubMedGoogle Scholar
- Ryan MAK, Gray GC, Smith B, McKeehan JA, Hawksworth AW, Malasig MD. Large epidemic of respiratory illness from adenovirus types 7 and 3 in healthy young adults. Clin Infect Dis. 2002. In press.
- McNeill KM, Benton FR, Monteith SC, Tuchscherer MA, Gaydos CA. Epidemic spread of adenovirus type 4-associated acute respiratory disease between U.S. army installations. Emerg Infect Dis. 2000;6:415–9.PubMedGoogle Scholar
- Li QG, Wadell G. Analysis of 15 different genome types of adenovirus type 7 isolated on five continents. J Virol. 1986;60:331–5.PubMedGoogle Scholar
- Li QG, Zheng QJ, Liu YH, Wadell G. Molecular epidemiology of adenovirus types 3 and 7 isolated from children with pneumonia in Beijing. J Med Virol. 1996;49:170–7. DOIPubMedGoogle Scholar
- Wadell G, Varsanyi TM. Demonstration of three different subtypes of adenovirus type 7 by DNA restriction site mapping. Infect Immun. 1978;21:238–46.PubMedGoogle Scholar
- Crawford-Miksza LK, Nang RN, Schnurr DP. Strain variation in adenovirus serotypes 4 and 7a causing acute respiratory disease. J Clin Microbiol. 1999;37:1107–12.PubMedGoogle Scholar
- Wadell G, Cooney MK, da Costa Linhares A, de Silva L, Kennett ML, Kono R, Molecular epidemiology of adenoviruses: global distribution of adenovirus 7 genome types. J Clin Microbiol. 1985;21:403–8.PubMedGoogle Scholar
- Adrian T, Becker M, Hierholzer JC, Wigand R. Molecular epidemiology and restriction site mapping of adenovirus 7 genome types. Arch Virol. 1989;106:73–84. DOIPubMedGoogle Scholar
- de Silva LM, Colditz P, Wadell G. Adenovirus type 7 infections in children in New South Wales, Australia. J Med Virol. 1989;29:28–32. DOIPubMedGoogle Scholar
- Moraes MT, Leite JP, Siqueira MM, Portes SA, Krawczuk MM, Nascimento JP. Genomic characterization of adenovirus serotype 7 isolated in Brazil from acute respiratory disease patients during the period from 1980 to 1991. Rev Inst Med Trop Sao Paulo. 1997;39:185–9. DOIPubMedGoogle Scholar
- Golovina GI, Zolotaryov FN, Yurlova TI. Sensitive analysis of genetic heterogeneity of adenovirus types 3 and 7 in the Soviet Union. J Clin Microbiol. 1991;29:2313–21.PubMedGoogle Scholar
- Kajon A, Wadell G. Genome analysis of South American adenovirus strains of serotype 7 collected over a 7-year period. J Clin Microbiol. 1994;32:2321–3.PubMedGoogle Scholar
- Kajon AE, Mistchenko AS, Videla C, Hortal M, Wadell G, Avendano LF. Molecular epidemiology of adenovirus acute lower respiratory infections of children in the south cone of South America (1991-1994). J Med Virol. 1996;48:151–6. DOIPubMedGoogle Scholar
- Larranaga C, Kajon A, Villagra E, Avendano LF. Adenovirus surveillance on children hospitalized for acute lower respiratory infections in Chile (1988-1996). J Med Virol. 2000;60:342–6. DOIPubMedGoogle Scholar
- Noda M, Kiritani M, Abe K, Ikeda Y, Yamaoka K, Ogino T. Restriction enzyme analysis of adenovirus type 7 isolates in Japan. Infectious agents surveillance report. National Institute of Infectious Diseases, Japan. 1996;17:102–4.
- Ikeda Y, Abe K, Kamimura M, Fujii A, Yamaoka K, Ogino T. Isolation of adenovirus type 7 during 1995-1999 and a case of type 7h infection - Hiroshima City. Infectious agents surveillance report. National Institute of Infectious Diseases, Japan. 2000;21:27–8.
- Azar R, Varsano N, Mileguir F, Mendelson E. Molecular epidemiology of adenovirus type 7 in Israel: identification of two new genome types, Ad7k and Ad7d2. J Med Virol. 1998;54:291–9. DOIPubMedGoogle Scholar
- Hashido M, Mukouyama A, Sakae K, Tsuzuki H, Yamashita T, Inada T, Molecular and serological characterization of adenovirus genome type 7h isolated in Japan. Epidemiol Infect. 1999;122:281–6. DOIPubMedGoogle Scholar
- Xu W, Erdman DD. Identification of human adenovirus types 3, 7 and 21 by a multiplex PCR assay. J Med Virol. 2001;64:537–42. DOIPubMedGoogle Scholar
- Deryckere F, Burgert HG. Rapid method for preparing adenovirus DNA. Biotechniques. 1997;22:868–70.PubMedGoogle Scholar
- Niel C, Moraes MT, Mistchenko AS, Leite JP, Gomes SA. Restriction site mapping of four genome types of adenovirus types 3 and 7 isolated in South America. J Med Virol. 1991;33:123–7. DOIPubMedGoogle Scholar
- Kajon AE, Wadell G. Characterization of adenovirus genome type 7h: analysis of its relationship to other members of serotype 7. Intervirology. 1992;33:86–90.PubMedGoogle Scholar
- Crawford-Miksza L, Schnurr DP. Analysis of 15 adenovirus hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues. J Virol. 1996;70:1836–44.PubMedGoogle Scholar
- Kajon AE, Murtagh P, Garcia Franco S, Freire MC, Weissenbacher MC, Zorzopulos J. A new genome type of adenovirus 3 associated with severe lower acute respiratory infection in children. J Med Virol. 1990;30:73–6. DOIPubMedGoogle Scholar
- Li Q, Wadell G. Genetic variability of hexon loops 1 and 2 between seven genome types of adenovirus serotype 7. Arch Virol. 1999;144:1739–49. DOIPubMedGoogle Scholar
- Fu WY, Dong L, Zheng YC, Liu WM, Zhong X, Guo HJ. A study of molecular epidemiology of adenovirus of types 3 and 7 on infant pneumonia in Northern China. Chin Med J (Engl). 1989;102:857–61.PubMedGoogle Scholar
- Chany C, Lepine P, Lelong M, Vin L-T, Stage P. Severe and fatal pneumonia in infants and young children associated with adenovirus infections. Am J Hyg. 1958;67:367–78.PubMedGoogle Scholar
- Ozawa S, Yamagami T, Watanabe T, Machida A, Yakutai K, Yokoyama H. Serosurvey after an outbreak of adenovirus type 7 infection at a high school dormitory - Yamanashi. Infectious agents surveillance report. National Institute of Infectious Diseases, Japan. 1997;18:82–3.
- Miwa C. [Serosurvey for adenovirus type 3 and adenovirus type 7 infection in resident in Gifu Prefecture, 1996]. Kansenshogaku Zasshi. 1997;71:1261–2.PubMedGoogle Scholar
- Ludwig SL, Brundage JF, Kelley PW, Nang R, Towle C, Schnurr DP, Prevalence of antibodies to adenovirus serotypes 4 and 7 among unimmunized US Army trainees: results of a retrospective nationwide seroprevalence survey. J Infect Dis. 1998;178:1776–8. DOIPubMedGoogle Scholar
- Piedra PA, Poveda GA, Ramsey B, McCoy K, Hiatt P. Incidence and prevalence of neutralizing antibodies to the common adenoviruses in children with cystic fibrosis: implication for gene therapy with adenovirus vectors. Pediatrics. 1998;101:1013–9. DOIPubMedGoogle Scholar
- Videla C, Carballal G, Misirlian A, Aguilar M. Acute lower respiratory infections due to respiratory syncytial virus and adenovirus among hospitalized children from Argentina. Clin Diagn Virol. 1998;10:17–23. DOIPubMedGoogle Scholar
Figure
Tables
Cite This Article1These data were presented in part at the International Conference on Emerging Infectious Diseases, July 16-19, 2000, Atlanta, GA.
Table of Contents – Volume 8, Number 3—March 2002
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Dean D. Erdman, Respiratory and Enteric Viruses Branch, Division of Viral and Rickettsial Diseases, Centers for Disease Control and Prevention, Mailstop G-09, Atlanta, GA 30333, USA; fax: 404-639-1307;
Top