Volume 12, Number 1—January 2006
Dispatch
Novel Parvovirus and Related Variant in Human Plasma
Cite This Article
Citation for Media
Abstract
We report a novel parvovirus (PARV4) and related variants in pooled human plasma used in the manufacture of plasma-derived medical products. Viral DNA was detected by using highly selective polymerase chain reaction assays; 5% of pools tested positive, and amounts of DNA ranged from <500 copies/mL to >106 copies/mL plasma.
Using a sequence-independent polymerase chain reaction (PCR) amplification method, we recently identified a new parvovirus in plasma from a patient with exposures and symptoms consistent with acute HIV infection, but who was HIV RNA negative (1). Phylogenetic analyses of sequence data suggest that this virus, termed PARV4, is only distantly related to previously known human or animal members of the family Parvoviridae, including members of the Erythrovirus genus known to infect humans, such as parvovirus B19. Infection with parvovirus B19, although frequently asymptomatic, may result in erythema infectiosum, arthropathy, pregnancy complications (e.g., hydrops fetalis), transient aplastic crisis, and disease in immunocompromised patients (2). Parvovirus B19 is most frequently transmitted through the respiratory route or vertically from mother to fetus. However, blood- and plasma-derived medical products, particularly clotting factors, contaminated with parvovirus B19 can also transmit the virus (3). Manufacturers of plasma derivatives screen minipools by using nucleic acid amplification techniques (NAT), which has enabled levels of erythrovirus DNA to be substantially reduced in start pools; for certain products, screening is now a regulatory requirement (4). This study examined pooled human plasma for fractionation to detect PARV4 DNA sequences.
Samples of manufacturing plasma pools submitted to the National Institute for Biological Standards and Control for testing for hepatitis C virus RNA were stored at –70°C until analysis, in compliance with European regulatory requirements. Manufacturing pools were sourced from donations collected in Europe and North America and received during the previous 6 months. Total nucleic acid was extracted from plasma pools as described previously (4) before analysis for PARV4 DNA.
Figure 1
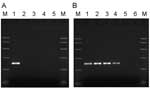
Figure 1. A) Specificity of primers for PARV4. Samples in lanes 1–5 were amplified by using primers directed to open reading frame 1 (ORF1) of PARV4. Template DNA in lane 1 was a...
Figure 2
![Thumbnail of Phylogenetic analysis of a 178-bp sequence of ORF1 of PARV4 and PARV5 (GenBank accession no. DQ112361) with other members of the Parvoviridae subfamily. The alignment includes the members of the Erythrovirus genus (parvovirus B19 [5]) and related viruses such as V9 (6), D91.1 (7), and A6 (8), as well as the closely related viruses infecting the cynomolgus macaque (LTMPV) (9) and rhesus (RMPV) and pig-tailed macaques (PTMPV) (10). Two other viruses tentatively assigned to the group i](/eid/images/05-0916-F2-tn.gif)
Figure 2. hylogenetic analysis of a 178-bp sequence of ORF1 of PARV4 and PARV5 (GenBank accession no. DQ112361) with other members of the Parvoviridae subfamily. The alignment includes the members of the Erythrovirus...
Using multiple sequence alignments of human erythroviruses and comparison with the sequence for PARV4 (1), we designed highly selective primers to the open reading frame 1 (ORF1) of PARV4, homologous to the nonstructural proteins of other parvoviruses. Primers PV4ORF1F (5´-AAGACTACATACCTACCTGTG-3´) and PV4ORF1R (5´-GTGCCTTTCATATTCAGTTCC-3´) amplify a 220-bp region of ORF1. The specificity of these primers was confirmed by PCR using a cloned fragment of the ORF1 region alongside erythrovirus control material (Figure 1A). Each PCR contained 1× PCR buffer II (PE Applied Biosystems, Warrington, UK), 200 μmol/L each deoxynucleoside triphosphate, 2 mmol/L MgCl2, 10 pmol each primer, and 2.5 U AmpliTaq Gold DNA polymerase (PE Applied Biosystems) in a final volume of 50 μL. For thermal cycling, a T3 thermal cycler (Biometra, Göttingen, Germany) was used with the following cycling conditions: 95°C for 9 min, followed by 45 cycles of 96°C for 30 s, 55°C for 30 s, and 72°C for 1 min. Amplicons were analyzed by agarose gel electrophoresis and compared to known size markers. The PARV4 control sequences (nucleotides 1293–1833 of ORF1, GenBank accession no. AY622943) were cloned into the vector pT7 Blue according to the manufacturer's instructions (Novagen, Darmstadt, Germany). The sensitivity of these PCR reactions was 1–10 copies of PARV4 sequences. DNA extracted from 137 pools was screened for PARV4 ORF1 sequences by PCR using 5 μL extracted DNA. Results, summarized in Table 1, show that 7 of 137 plasma pools screened with these primers tested positive for PARV4 DNA sequences and those of a related variant, known as PARV5. Typical results from pools and control plasmid samples are shown in Figure 1B. DNA sequence analysis showed that PARV5, over the region amplified, shares ≈92% nucleotide identity with PARV4 (Figure 2). Further sequence analysis around the primer-binding sites showed that the primers were 100% homologous in both genotypes. This level of relatedness is similar to that seen for the different erythrovirus genotypes (7).
The levels of PARV4 in the positive plasma pools were determined by real-time PCR using the screening primers from the ORF1 region of PARV4. Amplification reactions were performed on the LightCycler instrument using the LightCycler FastStart DNA MasterPLUS SYBR green I kit (Roche Applied Science, Mannheim, Germany) in accordance with the manufacturers' instructions. A standard curve was generated from the cloned plasmid DNA containing the ORF1 fragment of PARV4. Levels of PARV4 DNA were as high as 3.9 × 106 copies/mL plasma, although several pools contained <500 copies/mL plasma (Table 2).
Plasma pools found positive for PARV4 sequences were tested for the levels of erythrovirus DNA as described previously (4). Only 2 of the PARV4-positive pools contained any human erythrovirus DNA, and these were at low levels (Table 2). Of the plasma pools found to be positive for PARV4 sequences, blood products from only 2 were available for further analysis. Both products were immunoglobulin preparations, and in neither case could PARV4 sequences be detected.
This report is the first to describe novel parvovirus sequences in pooled human plasma for fractionation. PARV4 was originally identified in a patient with acute viral infection syndrome coinfected with hepatitis B virus (1). As yet, nothing is known about the prevalence of PARV4, its possible role in human disease, or whether PARV4 was transmitted to the original patient from an unidentified animal host.
Although PARV4 shares limited homology with human erythroviruses, the latter are frequent contaminants of plasma, pooled and used for fractionation (3). Levels of PARV4 DNA ranged from <500 copies/mL to >106 copies/mL plasma. If a single donation with a high PARV4 count was responsible for the contamination of such a pool, the levels of virus DNA in the original donation would have been in the order of 109 or 1010 copies/mL plasma, given the volume of the start pool. Because erythroviruses are small, nonenveloped, and relatively resistant to virus inactivation procedures, manufacturers of plasma-derived products have used NAT to exclude high-titer donations from manufacturing start pools. Before such measures were introduced, more than half of production start pools contained erythrovirus DNA, some with titers of 109 copies/mL plasma (4; S. Baylis, unpub. data). The prevalence of PARV4 and PARV5 and the titers observed in the pools examined in this study are much lower than the usual prevalence and titers observed with erythroviruses. Because of PARV4's insufficient homology with human erythroviruses, current methods of NAT are unlikely to identify donations positive for PARV4.
The availability of highly specific reagents for PARV4 and PARV5 will assist in further studies to elucidate their possible role in human disease. The detection of PARV4 and PARV5 in plasma may have been caused by an epidemic at the time of plasma donation. In a recent study that screened for enteroviruses in human plasma, seasonal changes were observed in the frequency and level of viremia (15). Studies to examine the epidemiology of PARV4 and PARV5 infection will help address issues such as these.
In summary, PARV4, a novel parvovirus, and PARV5, a related variant, have been identified in plasma used in the manufacture of blood products. Plasma is obtained from healthy persons, who at the time of donation are asymptomatic, despite being viremic for PARV4 or PARV5. Highly specific and sensitive assays to detect PARV4 will facilitate further analysis of the role of this novel virus in human disease and the implications of virus transmission by contaminated blood and blood products.
After this article was submitted for publication, human bocavirus, a novel parvovirus, was identified in respiratory tract samples (16). PARV4 and PARV5 are distinct from human bocavirus. For example, comparison of PARV4 (AY622943) with human bocavirus strains ST and ST2 (DQ000495 and DQ000496) shows nucleotide identity of 41% and 40%, respectively.
Dr Fryer is a scientist at the National Institute for Biological Standards and Control. Her work focuses on the quality of blood and blood products with respect to transfusion-transmitted infections.
Acknowledgment
We thank Nita Shah for technical assistance.
References
- Jones MS, Kapoor A, Lukashov VV, Simmonds P, Hecht F, Delwart E. New DNA viruses identified in patients with acute viral infection syndrome. J Virol. 2005;79:8230–6. DOIPubMedGoogle Scholar
- Heegaard ED, Brown KE. Human parvovirus B19. Clin Microbiol Rev. 2002;15:485–505. DOIPubMedGoogle Scholar
- Laub R, Strengers P. Parvoviruses and blood products. Pathol Biol (Paris). 2002;50:339–48. DOIPubMedGoogle Scholar
- Baylis SA, Shah N, Minor PD. Evaluation of different assays for the detection of parvovirus B19 DNA in human plasma. J Virol Methods. 2004;121:7–16. DOIPubMedGoogle Scholar
- Shade RO, Blundell MC, Cotmore SF, Tattersall P, Astell CR. Nucleotide sequence and genome organization of human parvovirus B19 isolated from the serum of a child during aplastic crisis. J Virol. 1986;58:921–36.PubMedGoogle Scholar
- Nguyen QT, Sifer C, Schneider V, Allaume X, Servant A, Bernaudin F, Novel human erythrovirus associated with transient aplastic anemia. J Clin Microbiol. 1999;37:2483–7.PubMedGoogle Scholar
- Servant A, Laperche S, Lallemand F, Marinho V, De Saint Maur G, Meritet JF, Genetic diversity within human erythroviruses: identification of three genotypes. J Virol. 2002;76:9124–34. DOIPubMedGoogle Scholar
- Nguyen QT, Wong S, Heegaard ED, Brown KE. Identification and characterization of a second novel human erythrovirus variant, A6. Virology. 2002;301:374–80. DOIPubMedGoogle Scholar
- Brown KE, Green SW, O'Sullivan MG, Young NS. Cloning and sequencing of the simian parvovirus genome. Virology. 1995;210:314–22. DOIPubMedGoogle Scholar
- Green SW, Malkovska I, O'Sullivan MG, Brown KE. Rhesus and pig-tailed macaque parvoviruses: identification of two new members of the erythrovirus genus in monkeys. Virology. 2000;269:105–12. DOIPubMedGoogle Scholar
- Yoo BC, Lee DH, Park SM, Park JW, Kim CY, Lee HS, A novel parvovirus isolated from Manchurian chipmunks. Virology. 1999;253:250–8. DOIPubMedGoogle Scholar
- Allander T, Emerson SU, Engle RE, Purcell RH, Bukh J. A virus discovery method incorporating DNase treatment and its application to the identification of two bovine parvovirus species. Proc Natl Acad Sci U S A. 2001;98:11609–14. DOIPubMedGoogle Scholar
- Hijikata M, Abe K, Win KM, Shimizu YK, Keicho N, Yoshikura H. Identification of new parvovirus DNA sequence in swine sera from Myanmar. Jpn J Infect Dis. 2001;54:244–5.PubMedGoogle Scholar
- Thompson JD, Higgins DG, Gibson TJ. ClustalW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–80. DOIPubMedGoogle Scholar
- Welch J, Maclaren K, Jordan T, Simmonds P. Frequency, viral loads, and serotype identification of enterovirus infections in Scottish blood donors. Transfusion. 2003;43:1060–6. DOIPubMedGoogle Scholar
- Allander T, Tammi MT, Eriksson M, Bjerkner A, Tiveljung-Lindell A, Andersson B. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proc Natl Acad Sci U S A. 2005;102:12891–6. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 12, Number 1—January 2006
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Sally A. Baylis, Division of Virology, National Institute for Biological Standards and Control, Blanche Lane, South Mimms, Potters Bar, Hertfordshire, EN6 3QG, UK; fax: 44-1707-646-730
Top