Volume 28, Number 9—September 2022
Dispatch
Sporadic Occurrence of Enteroaggregative Shiga Toxin–Producing Escherichia coli O104:H4 Similar to 2011 Outbreak Strain
Cite This Article
Citation for Media
Abstract
We describe the recent detection of 3 Shiga toxin–producing enteroaggregative Escherichia coli O104:H4 isolates from patients and 1 from pork in the Netherlands that were genetically highly similar to isolates from the 2011 large-scale outbreak in Europe. Our findings stress the importance of safeguarding food supply production chains to prevent future outbreaks.
Shiga toxin–producing Escherichia coli (STEC) is a zoonotic pathogen that causes illness ranging from mild diarrhea to hemolytic uremic syndrome and death. During 2011, an exceptionally large outbreak caused by serotype O104:H4 STEC occurred in Europe, mainly in Germany and France, that was associated with sprouts grown from imported fenugreek seeds (1). Besides the ability to produce Shiga toxin, specifically stx2a, the strain had the genetic characteristics and phylogenetic backbone of an enteroaggregative E. coli (EAEC) pathotype (2) but lacked other classical STEC virulence markers eae and hlyA (3). In addition, the outbreak strain carried plasmid-borne blaCTX-M-15 and blaTEM-1 genes. The epidemiologic investigation revealed that a contaminated batch of fenugreek seeds imported into the European Union from Egypt was the most probable source of the pathogen causing the outbreak (4).
After the 2011 outbreak in Germany and France, only a few sporadic cases of infection with Shiga toxin–producing EAEC O104:H4 were reported, most related to travel to Turkey or North Africa (5–8). We describe the sporadic occurrence of Shiga toxin–producing EAEC O104:H4 isolates in the Netherlands, originating from 2 clinical cases from 2019 and 2020 and 1 food isolate from 2017. In addition, we report a clinical case from Austria in 2021.
Surveillance of Shiga toxin–producing E. coli (STEC) in the Netherlands is performed in 1 or both of 2 ways: by mandatory case notification from medical laboratories or physicians to regional public health services, which then report the cases to the Netherlands National Institute for Public Health and the Environment (RIVM), or by microbiological surveillance, when medical laboratories send suspected STEC isolates to RIVM for confirmation and further molecular characterization. Since 2016, STEC isolates have been subjected to whole-genome sequencing for high-resolution detection of clusters and outbreaks nationwide. During detailed retrospective analysis of the whole-genome sequencing data, we identified isolates from 2 clinical cases of STEC infection from 2019 and 2020 and characterized them as stx2a-encoding EAEC O104:H4 (isolates NL1981 and NL2076). Both cases involved middle-aged nonhospitalized women with abdominal cramps and bloody diarrhea. Neither of the 2 case-patients reported recent travel. The 2019 case-patient reported consuming cooked minced beef, and the 2020 case-patient reported consuming beef, hamburger meat, and vegetables and fruits from her own garden. In 2021, the Austrian Agency for Health and Food Safety obtained a sample of similar stx2a-encoding EAEC O104:H4 from a 10-year-old girl with hemolytic uremic syndrome. This case-patient also reported no recent travel, but she had consumed raw veal. Finally, we retrospectively identified stx2a-encoding EAEC O104:H4 in a sample obtained from pig meat (pork) in 2017 in the Netherlands.
Figure 1

Figure 1. Single-linkage hierarchical clustering tree of enteroaggregative Shiga toxin–producing Escherichia coliO104:H4 from the Netherlands and reference sequences. Tree results from core- and accessory-genome multilocus sequence typing with a heatmap...
Figure 2
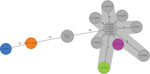
Figure 2. Minimum-spanning tree from cgMLST (Enterobase STEC scheme) of enteroaggregative Shiga toxin–producing Escherichia coliO104:H4 from the Netherlands and reference sequences. Colored isolates are those added during this study: green...
We performed a genomic comparison among the recently identified isolates, 2 Shiga toxin–producing EAEC O104:H4 isolates in cases from the Netherlands from 2013 described elsewhere (8), and a set of representative isolates from a 2011 outbreak in Germany and France, including isolates from case-patients in Germany who shed the outbreak strain well into 2012 (long-term shedders). To achieve our full set of isolates (Table), we supplemented the dataset with O104:H4 isolates retrieved from public repositories to gain more insights into the relatedness among investigated isolates and track possible evolutionary events. We assessed genomic relations between isolates by core- and accessory-genome multilocus sequence typing using the Enterobase STEC schema implemented in Ridom Seqsphere version 8.0.1 (https://www.ridom.de). The resulting dendrogram was rooted with strain 1060_13, a Clermont type A isolate. The 2020 patient isolate from the Netherlands (NL2076) and the pork isolate (ESBL3427) clearly clustered with the representative outbreak isolates (Figures 1,2). Conversely, patient isolates NL1981 from the Netherlands in 2019 and MRV-21-00239 from Austria in 2021 appeared more similar to each other and the 2 patient isolates (338 and 381_1) from the Netherlands in 2013 than to isolates from the outbreak cluster (Figures 1,2).
We determined the content of the plasmid, virulence, and antimicrobial resistance genes using the PlasmidFinder, VirulenceFinder, and ResFinder databases (http://www.genomicepidemiology.org). Within the outbreak cluster, including the patient (NL2076) and pork (ESBL3427) strains from the Netherlands, isolates showed virulence profiles typical of the 2011 outbreak strain, including STEC virulence markers stx2a, lpfA, and iha; the EAEC pAA plasmid (with aggR, aar, aap, sepA, the aatPABCD operon, and the aggABCD operon); and some chromosomal EAEC markers such as pic and sigA, but they lacked the STEC hallmark intimin eae. In addition, the isolates within the outbreak cluster showed similar antimicrobial resistance profiles, including blaCTX-M-15 and blaTEM-1B. Exceptions were the 2012 isolates from long-term shedders in Germany, which lost the pAA plasmid but belong to the outbreak cluster on the basis of their core genome (9) (Figure 1). Ancestral trait reconstruction as implemented in the R package ape v.5.5 (10), indicated a high likelihood that the most recent common ancestor of the isolates identified in this study were lysogenized with an stx-encoding phage. Among the isolates in the outbreak cluster, the core-genome allelic distance was 0–15 alleles (Figure 2).
After the 2011 outbreak in Europe, only a few sporadic cases of infection with stx-producing EAEC O104:H4 strains, most related to travel to Turkey or North Africa (8,11), were reported. The fact that the particular EAEC O104:H4 strains have never convincingly been isolated from food, animal, or environmental sources, and that EAEC in general are primarily human-adapted (12), support the hypothesis of a human reservoir and potential multiple events of import by travelers from an area where this pathogen is endemic. On the other hand, the loss of essential virulence markers in long-term shedders might be a preview of common trends of bacterial genome reduction in long-term carriage in humans which, in turn, might indicate that the newly signaled isolates are maintained in niches other than the human one. Genomic analysis of several post-outbreak EAEC O104:H4 isolates suggests that they are not derived from the 2011 outbreak but share a recent common ancestor (13). Our analysis indicates that distinct more- or less-distant variants of Shiga toxin–producing EAEC O104:H4 are circulating worldwide. These variants are more likely to represent independent evolutionary events than continuous diversification of a single clade established and circulating in Europe after the large 2011 outbreak (14). In the absence of any previous indication of Shiga toxin–producing EAEC O104:H4 in animals, it was surprising to retrieve such an isolate from a pork product. However, this finding does not necessarily imply that pigs are a reservoir, because the contamination could also have originated from a food handler or contaminated feed rather than the pigs.
In conclusion, we show that Shiga toxin–producing EAEC O104:H4 isolates highly related to the 2011 outbreak strain are sporadically occurring in Europe. We emphasize the need to optimize safeguarding vulnerable chains of food production to prevent future outbreaks.
Dr. Coipan is a genomic epidemiologist in the department of epidemiology and surveillance of enteric infections and zoonoses. Her primary research focus is the integration of bioinformatics, microbial ecology, population genetics, and classical epidemiology to understand origins and spread of pathogens.
Acknowledgment
The authors thank GGD, the Dutch municipal health services, for interviewing the STEC case-patients and the MML medical laboratories for submitting STEC isolates for additional typing. We thank Kees Veldman for his assistance in tracking the food strain history, reculturing the strain for whole genome sequencing, and reviewing the manuscript.
References
- Buchholz U, Bernard H, Werber D, Böhmer MM, Remschmidt C, Wilking H, et al. German outbreak of Escherichia coli O104:H4 associated with sprouts. N Engl J Med. 2011;365:1763–70. DOIPubMedGoogle Scholar
- Scheutz F, Nielsen EM, Frimodt-Møller J, Boisen N, Morabito S, Tozzoli R, et al. Characteristics of the enteroaggregative Shiga toxin/verotoxin-producing Escherichia coli O104:H4 strain causing the outbreak of haemolytic uraemic syndrome in Germany, May to June 2011. Euro Surveill. 2011;16:19889. DOIPubMedGoogle Scholar
- Bielaszewska M, Mellmann A, Zhang W, Köck R, Fruth A, Bauwens A, et al. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: a microbiological study. Lancet Infect Dis. 2011;11:671–6. DOIPubMedGoogle Scholar
- EFSA. Shiga toxin-producing E. coli (STEC) O104:H4 2011 outbreaks in Europe: taking stock. EFSA J. 2011;9:2390. DOIGoogle Scholar
- Jourdan-da Silva N, Watrin M, Weill FX, King LA, Gouali M, Mailles A, et al. Outbreak of haemolytic uraemic syndrome due to Shiga toxin-producing Escherichia coli O104:H4 among French tourists returning from Turkey, September 2011. Euro Surveill. 2012;17:20065. DOIPubMedGoogle Scholar
- Frank C, Milde-Busch A, Werber D. Results of surveillance for infections with Shiga toxin-producing Escherichia coli (STEC) of serotype O104:H4 after the large outbreak in Germany, July to December 2011. Euro Surveill. 2014;19:20760. DOIPubMedGoogle Scholar
- Guy L, Jernberg C, Arvén Norling J, Ivarsson S, Hedenström I, Melefors Ö, et al. Adaptive mutations and replacements of virulence traits in the Escherichia coli O104:H4 outbreak population. PLoS One. 2013;8:
e63027 . DOIPubMedGoogle Scholar - Ferdous M, Zhou K, de Boer RF, Friedrich AW, Kooistra-Smid AM, Rossen JW. Comprehensive characterization of Escherichia coli O104:H4 isolated from patients in the Netherlands. Front Microbiol. 2015;6:1348. DOIPubMedGoogle Scholar
- Zhang W, Bielaszewska M, Kunsmann L, Mellmann A, Bauwens A, Köck R, et al. Lability of the pAA virulence plasmid in Escherichia coli O104:H4: implications for virulence in humans. PLoS One. 2013;8:
e66717 . DOIPubMedGoogle Scholar - Paradis E, Schliep K. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics. 2019;35:526–8. DOIPubMedGoogle Scholar
- Grad YH, Godfrey P, Cerquiera GC, Mariani-Kurkdjian P, Gouali M, Bingen E, et al. Comparative genomics of recent Shiga toxin-producing Escherichia coli O104:H4: short-term evolution of an emerging pathogen. MBio. 2013;4:e00452–12. DOIPubMedGoogle Scholar
- Okeke IN, Wallace-Gadsden F, Simons HR, Matthews N, Labar AS, Hwang J, et al. Multi-locus sequence typing of enteroaggregative Escherichia coli isolates from Nigerian children uncovers multiple lineages. PLoS One. 2010;5:
e14093 . DOIPubMedGoogle Scholar - Grad YH, Lipsitch M, Feldgarden M, Arachchi HM, Cerqueira GC, Fitzgerald M, et al. Genomic epidemiology of the Escherichia coli O104:H4 outbreaks in Europe, 2011. Proc Natl Acad Sci U S A. 2012;109:3065–70. DOIPubMedGoogle Scholar
- Tietze E, Dabrowski PW, Prager R, Radonic A, Fruth A, Auraß P, et al. Comparative genomic analysis of two novel sporadic Shiga toxin-producing Escherichia coli O104:H4 strains isolated 2011 in Germany. PLoS One. 2015;10:
e0122074 . DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: August 11, 2022
Table of Contents – Volume 28, Number 9—September 2022
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Eelco Franz, Antonie van Leeuwenhoeklaan 9, 3721 MA Bilthoven, Netherlands
Top