Volume 3, Number 4—December 1997
Dispatch
Vero Cytotoxin-Producing Escherichia coli O157 Outbreaks in England and Wales, 1995: Phenotypic Methods and Genotypic Subtyping
Cite This Article
Citation for Media
Abstract
Vero cytotoxin-producing Escherichia coli O157 belonging to four phage types (PTs) caused 11 outbreaks of infection in England and Wales in 1995. Outbreak strains of different PTs were distinguishable by DNA-based methods. Pulsed-field gel electrophoresis best discriminated among strains belonging to the same PT, distinguishing six of the seven PT2 outbreak strains and both PT49 outbreak strains.
Vero cytotoxin-producing Escherichia coli O157 (O157 VTEC), first associated with outbreaks of human disease in North America in 1982, has since emerged as an important human pathogen; it causes mild nonbloody diarrhea, hemorrhagic colitis, and hemolytic uremic syndrome, as well as less common manifestations such as thrombotic thrombocytopenic purpura with neurologic symptoms (1). Laboratory-confirmed infections with O157 VTEC in England and Wales increased from fewer than 10 in 1983 to 250 in 1990, 792 in 1995, and 660 in 1996 (2;3; Laboratory of Enteric Pathogens, unpub. data). The incidence of O157 VTEC infection in Scotland relative to its population is up to six times that in England and Wales, but wide geographic variations exist in England, Wales, and Scotland (4,5). The main reservoir for O157 VTEC appears to be healthy cattle, although recently organisms have been found in sheep (4,6,7). O157 VTEC infections are usually foodborne, associated with consumption of undercooked minced beef (most commonly as beefburgers), unpasteurized milk, and a variety of other vehicles such as salami, cheese, yogurt, water, salad vegetables, and fruit juice (7,8). Other routes of infection include animal contact and person-to-person spread, both in families and institutional settings (4).
O157 VTEC is differentiated by phage typing into more than 80 phage types (PTs) (9, R. Khakhria, pers. comm.). Polynucleotide DNA probes identify Vero cytotoxin (VT) genotypes and divide O157 VTEC into strains with VT1, VT2, or VT1+2 genes. However, certain phage and VT types predominate: thus in England and Wales approximately 50% of O157 VTEC are PT2 and produce VT2 (2,3). A range of DNA-based methods is available for further strain discrimination (4), and we have applied some of these to compare strains associated with outbreaks of O157 VTEC infection in England and Wales in 1995.
Eleven general outbreaks during 1995 affected members of more than one household or residents of an institution (Table), whereas 18 general outbreaks were reported during 1992 to 1994 (10). Most of the outbreaks in 1995 occurred in late summer and in the community (Table, outbreaks 5, 9, and 10); institutions (Table, outbreaks 1, 4, and 11); catering establishments (Table, outbreaks 3, 6, and 7); or a mixture of these (Table, outbreaks 2 and 8). Outbreaks occurred throughout England and Wales; of the 11, four were in the northern region. The incidence of hemolytic uremic syndrome was 0% to 36% in individual outbreaks and 8% overall. The case-fatality rate was 6%, mainly among the elderly. In six outbreaks, there was epidemiologic evidence for foodborne infection, but O157 VTEC was never isolated from food. Person-to-person spread was probably important in three outbreaks. The environmental and epidemiologic investigations of two of these incidents have been reported (11, 12).
For most outbreaks, epidemiologic investigation, phage typing of isolates, and further tests with VT1 and VT2 probes (Table) initially discriminated probable outbreak-associated cases from sporadic infections at the same time in the same area. Seven outbreaks were due to strains of PT2, VT2; two to PT49, VT2; and one to PT1, VT1+VT2. The other outbreak was associated with a VT1+VT2 strain that reacted with the typing phages but did not conform to a recognized type (reacts but does not conform [RDNC]). In outbreak 8, the O157 VTEC was resistant to sulphonamides and tetracyclines, whereas the other outbreak strains were sensitive to antimicrobial agents. In relation to community outbreaks 2, 5, and 8, several cases were infected with O157 VTEC that were similar to the outbreak strains, but the patients had no known epidemiologic link with the outbreak. Such strains were therefore included in the tests described below to provide evidence for the possible involvement of these cases. O157 VTEC that hybridize with the VT2 polynucleotide probe may carry different VT2 sequences (13). Strains from humans commonly possess VT2 or VT2c or both sequences. Polymerase chain reaction (14) showed that strains from all outbreaks except 2 and 3 possessed both types of VT2 sequences (Table). O157 VTEC belonging to PT1, such as those from outbreak 3, usually carry VT1 and only the VT2 sequence (13). The presence of the VT2, but not the VT2c, sequence differentiated the PT2 strain from outbreak 2 from all the other PT2-associated outbreaks. This property was exploited during the course of the outbreak to exclude patients infected with PT2 strains that had the VT2+VT2c genotype and not the outbreak VT2 genotype.
Figure
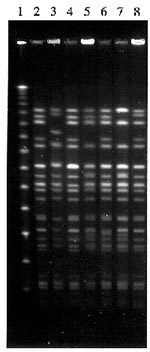
Figure. Pulsed-field gel electrophoresis of XbaI-digested genomic DNA of O157 VTEC PT2 isolated from outbreaks in 1995. Digests were separated on 1% agarose for 42h at a voltage gradient of 5.6 volts...
Pulsed-field gel electrophoresis (PFGE) has been applied widely as a highly discriminatory method for fingerprinting bacterial pathogens. The method of Barrett et al. (16), modified as indicated in the Figure, identified between 15 and 20 XbaI-generated fragments of the O157 VTEC strains (Figure). PFGE patterns of PT1, PT49, and RDNC were reproducible and clearly distinct from each other; they differed from the most common PT2 pattern by at least six fragment positions. The PT49 strains associated with outbreaks 7 and 11 were distinguished from each other by their PFGE profile. Results of PFGE of O157 VTEC of PT2 from the outbreaks are shown in the Figure. The strains that caused outbreaks 1 and 4 were indistinguishable by all methods including PFGE (lanes 2 and 4). Two other strains, from outbreaks 6 and 9, were very similar to these two, differing at only one fragment position (lanes 6 and 8), but the patterns were distinct and reproducible.
The PT2 strains from outbreaks 5 and 8 (lanes 5 and 7) differed from those associated with outbreaks 1 and 4 by at least 3 fragment positions and were distinguishable from each other. The strain from outbreak 2, which possessed only the VT2 gene, was clearly distinct from all the other PT2 strains by PFGE (lane 3). Criteria for the interpretation of patterns produced by PFGE have been published by Tenover et al. (17). Results of the analysis of O157 VTEC by PFGE suggest these criteria need modification for closely related organisms such as O157 VTEC (18).
Although phage typing and polynucleotide probes for VT1 and VT2 genes rapidly characterized strains from the outbreaks in 1995, DNA-based methods were valuable in distinguishing outbreak cases from outlying ones. VT2 gene subtyping was rapid but not very discriminatory, whereas RFLP and PFGE techniques differentiated strains but were time-consuming. The highest level of discrimination (by PFGE) distinguished certain O157 VTEC that appeared identical by all other methods. Some of the differences detected by PFGE were minor, but for each outbreak, they were reproducible with all the strains examined in this study. The epidemiologic or evolutionary significance of these minor variations is difficult to evaluate.
Investigation of the epidemiology of outbreaks of O157 VTEC infection requires a combined use of typing and fingerprinting methods in a hierarchic manner consistent with practical and economic constraints. In outbreaks well defined by epidemiologic studies, phage typing and the identification of VT1 and VT2 genes, including VT2 subtyping, are likely to be sufficient. In outbreaks less clearly defined epidemiologically, DNA-based methods may assist in identifying those strains not associated with the outbreak; this is particularly helpful when the outbreak is due to a common phage type. DNA-based methods have been useful in linking human infections with associated foods (15) and animals (19).
Although PFGE gives a high level of discrimination between closely related O157 VTEC, it has certain disadvantages. It is time-consuming and may not be suitable for rapid identification of large numbers of strains. We recommend the use of a combination of phage typing, VT typing, and PFGE to provide good discrimination of O157 VTEC strains in epidemiologic investigations.
Acknowledgment
We thank laboratories in England and Wales for sending clinical samples and bacterial isolates and colleagues in the Laboratory of Enteric Pathogens for technical assistance.
References
- Karmali MA. Infection by Verocytotoxin-producing Escherichia coli. Clin Microbiol Rev. 1989;2:15–38.PubMedGoogle Scholar
- Thomas A, Chart H, Cheasty T, Smith HR, Frost JA, Rowe B. Vero cytotoxin-producing Escherichia coli, particularly serogroup O157, associated with human infections in the United Kingdom: 1989 to 1991. Epidemiol Infect. 1993;110:591–600. DOIPubMedGoogle Scholar
- Thomas A, Cheasty T, Frost JA, Chart H, Smith HR, Rowe B. Vero cytotoxin-producing Escherichia coli, particularly serogroup O157, associated with human infections in England and Wales: 1992-4. Epidemiol Infect. 1996;117:1–10. DOIPubMedGoogle Scholar
- Advisory Committee on the Microbiological Safety of Food. Report on Verocytotoxin-producing Escherichia coli. London: HMSO; 1995.
- Sharp JCM, Reilly WJ, Coia JE, Curnow J, Synge BA. Escherichia coli O157 infection in Scotland: an epidemiological overview, 1984-94. PHLS Microbiology Digest. 1995;12:134–40.
- Chapman PA, Siddons CA, Harkin MA. Sheep as a potential source of verocytotoxin-producing Escherichia coli O157. Vet Rec. 1996;138:23–4.PubMedGoogle Scholar
- Griffin PM, Tauxe RV. The epidemiology of infections caused by Escherichia coli O157:H7, other enterohemorrhagic E. coli, and the associated hemolytic uremic syndrome. Epidemiol Rev. 1991;13:60–98.PubMedGoogle Scholar
- Neill MA, Tarr PI, Taylor DN, Trofa AF. Escherichia coli. In: Hui YH, Gorham JR, Murrell KD, Oliver DO, editors. Foodborne disease handbook 1994. New York: Marcel Dekker; 1994. p. 169-213.
- Khakhria R, Duck D, Lior H. Extended phage-typing scheme for Escherichia coli O157:H7. Epidemiol Infect. 1990;105:511–20. DOIPubMedGoogle Scholar
- Wall PG, McDonnell RJ, Adak GK, Cheasty T, Smith HR, Rowe B. General outbreaks of Vero cytotoxin-producing Escherichia coli O157 in England and Wales from 1992-1994. Commun Dis Rep CDR Rev. 1996;6:R26–33.PubMedGoogle Scholar
- Stevenson J, Hanson S. Outbreak of Escherichia coli O157 phage type 2 infection associated with eating precooked meats. Commun Dis Rep CDR Rev. 1996;6:R116–8.PubMedGoogle Scholar
- Gammie AJ, Mortimer PR, Hatch L, Brierley AFM, Chada N, Walters JB. Outbreak of Vero cytotoxin-producing Escherichia coli O157 associated with cooked ham from a single source. PHLS Microbiology Digest. 1996;13:142–5.
- Thomas A, Smith HR, Rowe B. Use of digoxigenin-labelled oligonucleotide DNA probes for VT2 and VT2 human variant genes to differentiate Vero cytotoxin-producing Escherichia coli strains of serogroup O157. J Clin Microbiol. 1993;31:1700–3.PubMedGoogle Scholar
- Thomas A, Cheasty T, Chart H, Rowe B. Isolation of Vero cytotoxin-producing Escherichia coli serotypes O9ab:H- and O101:H- carrying VT2 variant gene sequences from a patient with haemolytic uraemic syndrome. Eur J Microbiol Infect Dis. 1994;13:1074–6. DOIGoogle Scholar
- Willshaw GA, Thirlwell J, Jones AP, Parry S, Salmon RL, Hickey M. Vero cytotoxin-producing Escherichia coli O157 in beefburgers linked to an outbreak of diarrhoea, haemorrhagic colitis and haemolytic uraemic syndrome in Britain. Lett Appl Microbiol. 1994;19:304–7. DOIPubMedGoogle Scholar
- Barrett TJ, Lior H, Green JH, Khakhria R, Wells JG, Bell BP, Laboratory investigation of a multistate food-borne outbreak of Escherichia coli O157:H7 by using pulsed-field gel electrophoresis and phage typing. J Clin Microbiol 1994;32:3013-7. Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Swaminathan B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol 1995;33:2233-9. bender jb, hedberg cw, besser jm, boxrud dj, macdonald kl, osterholm mt. surveillance for Escherichia coli O157 infections in Minnesota by molecular subtyping. N Engl J Med. 1997;337:388–94. DOIPubMedGoogle Scholar
- Trevena WB, Willshaw GA, Cheasty T, Wray C, Gallagher J. Vero cytotoxin-producing E. coli O157 infection associated with farms. Lancet. 1996;347:60–1. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleTable of Contents – Volume 3, Number 4—December 1997
| EID Search Options |
|---|
|
|
|
|
|
|