Volume 31, Number 2—February 2025
Research Letter
Venezuelan Equine Encephalitis Virus Infection in Nonhuman Primate, Guatemala, 2023
Cite This Article
Citation for Media
Abstract
We isolated Venezuelan equine encephalitis virus (VEEV) subtype IE phylogenetically related to Gulf Coast strains in a spider monkey (Ateles geoffroyi) released from a rescue center in Guatemala. Serologic testing of 118 monkeys indicated no additional VEEV infections. Infection of a primate warrants intensified surveillance of VEEV transmission cycles in North America.
Venezuelan equine encephalitis virus (VEEV) is an alphavirus in the Americas that can cause febrile illness and severe disease, including encephalitis. In humans, overall case-fatality rates are <1% but higher in children; in horses, case-fatality rates are 50%–70% (https://www.woah.org/en/disease/venezuelan-equine-encephalitis). In the United States, VEEV is classified as a select agent because of its pathogenicity and aerosolization capacity (https://www.selectagents.gov/sat/list.htm). The transmission of the arthropodborne VEEV involves an epizootic cycle (antigenic subtypes IAB and IC), entailing higher numbers of human infections, and an enzootic cycle (the common transmission cycle for antigenic subtypes ID and IE), entailing sporadic human infections (1,2). The emergence of VEEV epizootics is poorly understood but might involve genetic exchanges from enzootic subtypes (1). VEEV subtype IE has been reported in Central America and Mexico since the 1960s (3). Subtype IE has been detected almost exclusively in mosquitoes and sentinel hamsters (3,4) but sporadically in horses and humans (2,3,5); the subtype has been associated with 2 epizootics in horses in Mexico in the 1990s (6). We report the detection and isolation of a VEEV IE strain in a healthy nonhuman primate (NHP) in Guatemala in 2023.
Figure 1
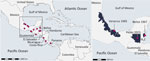
Figure 1. Geographic origins of reported Venezuelan equine encephalitis virus subtype IE sequences identified in Central America. Circles indicate geolocation of subtype IE sequences. Enlarged map shows the geolocation of the strains...
Figure 2

Figure 2. Time-scaled Bayesian maximum-clade credibility tree of Venezuelan equine encephalitis virus subtype IE identified in Central America. Bayesian phylogeny of the concatenated nonstructural and structural open reading frames with removal of...
We investigated 211 animals from 22 species (orders Artiodactyla, Carnivora, Didelphimorphia, Pilosa, Primates, and Rodentia) (Appendix Figure 1). We collected plasma samples from animals living in the Wildlife Rescue and Conservation Association (ARCAS) (https://arcasguatemala.org), a nongovernment organization in Petén Department in northern Guatemala. Animals are brought to ARCAS after being seized by police or customs officials at roadblocks or local markets. All animals are tested against selected pathogens and quarantined for 2–3 months upon arrival to the center and usually stay for another few months before being released into the wild (Appendix). We extracted RNA from plasma samples and screened it for alphaviruses and flaviviruses by using broadly reactive reverse transcription PCR (RT-PCR) assays (Appendix). By using an alphavirus RT-PCR, we identified VEEV in 1 adult male spider monkey (Ateles geoffroyi), an endangered species occurring from southern Mexico to Panama. No sample was positive for flavivirus RNA. Viral load in plasma was 1.8 × 105 VEEV RNA copies/mL, determined through real-time RT-PCR (Appendix). At 2 days postinfection, Vero E6 cells displayed a cytopathic effect and reached high concentrations of 1.4 × 1010 VEEV RNA copies/mL of supernatant. We applied deep sequencing (Appendix) to the virus isolate and obtained a near-complete genome (4.7 million reads, >50,000× mean depth of coverage), encompassing 11,477 nt and lacking only 12 nt in the 5′ untranslated region (GenBank accession no. PQ406672). The VEEV genome showed typical organization; the predicted regions encoded nonstructural protein genes (nsP1–nsP4) and structural protein genes (capsid, E1–E3, and 6K). The VEEV from Guatemala had pairwise nucleotide sequence distances of 1.4%–7.4% with VEEV subtype IE sequences available in GenBank that mostly originated from Central America (Figure 1). We identified no amino acid exchange at position 117 of the E2 glycoprotein, which previously had been associated with a VEEV subtype IE infection outbreak in horses (1). Pressure analyses that identify pervasive and episodic selection showed no sign of adaptive mutation in the E2 glycoprotein (Appendix Figure 2), suggesting strong purifying selection that might limit adaptation to new hosts (3). Bayesian phylogenetic analysis showed that the NHP-associated virus grouped with viruses obtained from the 1960s circulating in the Gulf Coast across a ≈200–700 km distance (most recent common ancestor 1954) (Figure 2), suggesting a lack of surveillance and continued circulation of genetically closely related strains in North America.
Natural VEEV infection in an NHP might be indicative of an outbreak. Therefore, we tested all plasma samples from primates residing in ARCAS (n = 118), including 80 spider monkeys and 38 howler monkeys (Alouatta pigra and A. palliata), for VEEV-specific IgM by using a modified commercial immunofluorescence assay (IFA) (Appendix). IFA detected no positive samples, including in the PCR-positive animal (Appendix Figure 3). The NHP infected with VEEV showed no clinical symptoms and was released into the wild on the day of blood sampling, suggesting that the animal was viremic upon release. In humans, severe disease characterized by neurologic complications occurs more frequently in children; therefore, future investigations might consider VEEV as a differential diagnosis, particularly in young NHP with acute neurologic disease in VEEV-endemic areas.
Although the neotropics are a probable hot spot for primate-associated emerging infections, neotropical NHPs are understudied for emerging pathogens (7). NHPs are among the most relevant sources of zoonotic viruses (in >20% of primate species zoonotic pathogens have been found) (7). Infection with other human pathogenic arthropodborne viruses, such as chikungunya and yellow fever viruses, has been reported in NHPs (8,9). Movement of NHPs by wildlife trafficking might contribute to the geographic expansion of VEEV and other pathogens. Although we identified no adaptive mutation in our study, the determinants of epizootics are not well understood, and new hosts might entail viral adaptation, potentially altering the viral phenotype. Seroepidemiologic and experimental infection studies, such as those conducted in rodents (10), are needed to clarify the role of NHPs in VEEV transmission cycles.
Dr. Jo is a virologist working as a postdoctoral researcher at the Institute of Virology at the Charité–Universitätsmedizin Berlin. Her research interests focus on the evolutionary origins of emerging viruses and the factors that drive their emergence.
Acknowledgments
We thank Melanie Roggow, Sabrina Figueroa, Kim Grützmacher, and Michael Nagel.
Export permits were approved by the local authority Consejo Nacional de Areas Protegidas (National Council for Protected Areas), including Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) permit nos. 493-494/2023, 496-497/2023, and 1363-64/2023, and non-CITES permit nos. 276/2023 and 398/2023. The export sanitary certificate (DPSP-E-15598) was approved by the Ministerio de Agricultura, Ganaderia, y Alimentacion (Minister of Agriculture and Food Supply) in Guatemala. Import permits were obtained from the Bundesamt für Naturschutz (Federal Agency for Nature Conservation), including CITES permit nos. DE-E-02185-86/23, DE-E-02188-86/23, DE-E05070/23, and DE-E05072/23, and import notification no. 223. Veterinary health permit nos. EG I-2023/60 and EG I-2024/08 were approved by the Senatsverwaltung für Justiz und Verbraucherschutz (Senate Department for Justice and Consumer Protection), in Berlin, Germany.
This work was supported by the German Federal Ministry of Economic Cooperation and Development through the Deutsche Gesellschaft für Internationale Zusammenarbeit GmbH (project no. 81284476), the European Union through the project Zoonosis Emergence across Degraded and Restored Forest Ecosystems (project no. 101135094), the Global Centers for Health and Pandemic Prevention (project no. 57592642), and a doctoral program scholarship from the German Academic Exchange Service (grant no. 91609043).
References
- Brault AC, Powers AM, Holmes EC, Woelk CH, Weaver SC. Positively charged amino acid substitutions in the e2 envelope glycoprotein are associated with the emergence of venezuelan equine encephalitis virus. J Virol. 2002;76:1718–30. DOIPubMedGoogle Scholar
- Scherer WF, Campillo-Sainz C, de Mucha-Macías J, Dickerman RW, Chia CW, Zarate ML. Ecologic studies of Venezuelan encephalitis virus in southeastern Mexico. VII. Infection of man. Am J Trop Med Hyg. 1972;21:79–85. DOIPubMedGoogle Scholar
- Forrester NL, Wertheim JO, Dugan VG, Auguste AJ, Lin D, Adams AP, et al. Evolution and spread of Venezuelan equine encephalitis complex alphavirus in the Americas. PLoS Negl Trop Dis. 2017;11:
e0005693 . DOIPubMedGoogle Scholar - Adams AP, Navarro-Lopez R, Ramirez-Aguilar FJ, Lopez-Gonzalez I, Leal G, Flores-Mayorga JM, et al. Venezuelan equine encephalitis virus activity in the Gulf Coast region of Mexico, 2003-2010. PLoS Negl Trop Dis. 2012;6:
e1875 . DOIPubMedGoogle Scholar - Aguilar PV, Estrada-Franco JG, Navarro-Lopez R, Ferro C, Haddow AD, Weaver SC. Endemic Venezuelan equine encephalitis in the Americas: hidden under the dengue umbrella. Future Virol. 2011;6:721–40. DOIPubMedGoogle Scholar
- Oberste MS, Fraire M, Navarro R, Zepeda C, Zarate ML, Ludwig GV, et al. Association of Venezuelan equine encephalitis virus subtype IE with two equine epizootics in Mexico. Am J Trop Med Hyg. 1998;59:100–7. DOIPubMedGoogle Scholar
- Han BA, Kramer AM, Drake JM. Global patterns of zoonotic disease in mammals. Trends Parasitol. 2016;32:565–77. DOIPubMedGoogle Scholar
- Althouse BM, Guerbois M, Cummings DAT, Diop OM, Faye O, Faye A, et al. Role of monkeys in the sylvatic cycle of chikungunya virus in Senegal. Nat Commun. 2018;9:1046. DOIPubMedGoogle Scholar
- Moreira-Soto A, Torres MC, Lima de Mendonça MC, Mares-Guia MA, Dos Santos Rodrigues CD, Fabri AA, et al. Evidence for multiple sylvatic transmission cycles during the 2016-2017 yellow fever virus outbreak, Brazil. Clin Microbiol Infect. 2018;24:1019.e1–4. DOIPubMedGoogle Scholar
- Deardorff ER, Forrester NL, Travassos-da-Rosa AP, Estrada-Franco JG, Navarro-Lopez R, Tesh RB, et al. Experimental infection of potential reservoir hosts with Venezuelan equine encephalitis virus, Mexico. Emerg Infect Dis. 2009;15:519–25. DOIPubMedGoogle Scholar
Figures
Cite This ArticleOriginal Publication Date: January 17, 2025
Table of Contents – Volume 31, Number 2—February 2025
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Jan Felix Drexler, Institute of Virology, Charitéplatz 1, 10117 Berlin, Germany
Top