Volume 32, Number 1—January 2026
Perspective
Emerging Respiratory Virus Threats from Influenza D and Canine Coronavirus HuPn-2018
Cite This Article
Citation for Media
Abstract
In 2009 and again in 2019, public health warnings were confirmed by the emergence, rapid widespread transmission, and lethality of novel influenza and coronaviruses. The world continues to suffer disease from these respiratory viruses. Two newly recognized emergent respiratory viruses, influenza D and canine coronavirus HuPn-2018, have been shown to have considerable potential for causing future human epidemics, but diagnostics and surveillance for the viruses are lacking. We reviewed data regarding influenza D virus and coronavirus canine coronavirus HuPn-2018. Those data strongly indicate that these viruses are major newly recognized threats. However, little is being done to respond to or prevent disease associated with these viruses, warranting the question of whether we will learn from previous pandemics.
Although science has developed effective countermeasures for most bacterial and vectorborne emerging pathogens, novel respiratory viruses continue to cause largescale human epidemics. Particularly problematic are pathogens that are of zoonotic origin. Viruses causing epidemics seem especially common among the Orthomyxoviridae and Coronaviridae viral families (1–6; https://www.who.int/emergencies/disease-outbreak-news/item/2025-DON560; https://data.who.int/dashboards/covid19/deaths?n=o) (Table). Those epidemics have routinely caught medical professionals off-guard and caused largescale disease and death. Two recently discovered viruses, influenza D and canine coronavirus HuPn-2018 (CCoV-HuPn-2018), seem especially worthy of closer public health attention.
Figure 1

Figure 1. Schematic illustration of the host range of IDV. Natural infections have been confirmed through the detection of virus genomic RNA, virus isolation, or the presence of IDV antibodies in domestic...
First detected and characterized in pigs with signs of respiratory illness in 2011, much has been learned about influenza D virus (IDV) since its first recognition (7,8). Like influenza A, B, and C viruses, IDVs are enveloped RNA viruses having segmented genomes that can change through reassortment, recombination, and mutation. IDVs belong to the genus Deltainfluenzavirus of the virus family Orthomyxoviridae. They share ≈50% amino acid identity with influenza C viruses (ICVs) across their genomes, but IDVs are much more prevalent in animal species. Initially thought to be enzootic in pigs and cattle, IDVs have now been detected in many livestock and wildlife species, including camels, deer, giraffes, kangaroos, llamas, wallabies, and wildebeests (9–14) (Figure 1). We have recently found evidence for IDV infections in poultry (12). A growing list of susceptible hosts for this new virus seems to be similar to those observed in the infection ecology of highly pathogenic avian influenza A(H5N1) viruses.
Like ICVs, a ubiquitous pathogen that causes minor influenza in humans, IDVs rely upon a hemagglutinin-esterase-fusion glycoprotein for cell binding and entry. Multiple recognized IDV strains or clades are chiefly classified by their hemagglutinin-esterase-fusion sequence circulating in animals. The genes of different IDV clades can reassort when they infect the same cell, and this mixing of genetic material leads to the generation of novel progeny viruses. Although science has had much more time to describe the ecology of ICVs, which were discovered in 1947, IDVs may reassort and recombine with other IDVs, suggesting that they are rapidly evolving (15). Although IDV prevalence is often high in cattle (G.C. Gray, unpub. data, 2025 Dec 11) compared with ICVs (16), IDVs may have more opportunity to gain characteristics that would threaten humans.
Although not all pig or cattle herds worldwide are affected by IDVs (17–19), many animal species clearly have infections periodically. The largest animal reservoir seems to be cattle, despite the first IDV being isolated from a diseased pig. The full spectrum of IDV illness in cattle is not known, but IDV is now recognized to contribute to one of the largest disease problems in cattle, bovine respiratory disease complex, which is estimated to cost the US cattle industry >$1 billion annually (20). As a measure of IDV endemicity, in our recent studies of 12 beef and dairy cattle farms in the United States and Mexico, we have detected (and often isolated) IDV >50 times among >500 nasal swab specimens obtained from sick or healthy cattle (G.C. Gray, unpub. data, 2025 Dec 11).
Although no viable (i.e., infectious) IDV has thus far been isolated from humans, mounting evidence indicates that the virus is zoonotic. Several human studies suggest that IDV causes subclinical infections in humans, especially among persons with occupational exposure to animals. In 2016, we reported a seroepidemiologic study of cattle workers in Florida, USA, where we found that >97% of cattle workers had neutralizing antibody to IDV, compared with 18% among a non–cattle exposed control population (13). In 2023, we reported a study of dairy workers in Colorado, where we found that 67% of 31 workers had molecular evidence of IDV in their nasal washes during a 5-day period (21).
Recently, a scientific team in China has reported compelling animal model, aerosol, and seroepidemiologic data that provide even stronger evidence that IDV is causing subclinical infections in humans (14). The team found that 73% of 612 study participants (97% among those with respiratory symptoms) in northeast China had serologic evidence of infection (14). They documented viral transmission in the air between ferrets, replication in primary human epithelial cells, infection in mice and dog models, and concluded that IDV has acquired the capacity for human-to-human transmission and that IDV strains already pose a potential panzootic threat (14). Of note, that study provides the first serologic evidence of widespread IDV in a general human population. The IDV strain in China that was used in the study, D/HY11, was isolated in 2023 from cattle and seems to be more efficient for airborne transmission in ferrets than strains used in previous studies. The observed increase in aerosol transmission is probably associated with mutations identified in this new strain, especially in the polymerase P3 gene, but more studies are needed to understand the underlying mechanisms behind these findings. Nevertheless, IDV replication and transmission in ferrets, 1 surrogate model for humans in influenza studies, and efficient replication in human primary airway epithelial cells observed in that work and another study (22) provide a theoretical framework that the virus will adapt and evolve for effective growth and human-to-human spread.
Figure 2
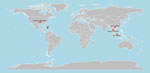
Figure 2. Locations of canine coronavirus HuPn-2018 or similar virus detections among humans with respiratory illness (23–27). Figure created in R version 4.4.1 (https://cran.rstudio.com).
In 2021, we first reported the cell culture isolation and characterization of a novel canine–feline recombinant alphacoronavirus, CCoV-HuPn-2018, from a nasopharyngeal swab sample from a child hospitalized with pneumonia in Sarawak State, Malaysia (23). The virus shared ≈97% nucleotide identity in most structural genes with canine coronavirus II, but its spike gene contained segments from feline coronavirus and transmissible gastroenteritis virus, which are specific for the canine coronavirus IIb subtype, suggesting a complex recombinant origin. A subsequent virus culture and characterization in urine specimens from persons visiting Haiti (24) indicated a 99.4% identity, confirming the circulation of CCoV-HuPn-2018 in different geographic regions. In addition, similar animal alphacoronaviruses have been detected among humans with respiratory illness living in Bangkok, Thailand (25), and in the US state of Arkansas (26). Recently, we detected CCoV-HuPN-2018 among 18 of 200 pneumonia patients hospitalized in the area of Hanoi, Vietnam, suggesting that this virus may have a wide geographic distribution and variable (and possibly increasing) prevalence (27) (Figure 2). The virus is entirely missed by common clinical diagnostics tests for the detection of respiratory viruses.
Although these findings do not yet prove that CCoV-HuPn-2018 is a frequent, worldwide cause of severe respiratory disease, they suggest that CCoV-HuPn-2018 (or very similar viruses) merit our closer study. Recent studies of the spike protein of CCoV-HuPN-2018 have shed light on its interaction with the aminopeptidase N from canines, felines, and porcines, but not humans, as functional receptors for cell entry (28–30). Nevertheless, CCoV-HuPn-2018 spike protein pseudotyped virus infects multiple human cancer cell lines in a human aminopeptidase N–independent manner. Earlier clades of CCoV-HuPn-2018 might have not yet evolved to be an efficient human pathogen, but they may be evolving now, as evidenced by the increased number of patients affected by the virus in the study by our surveillance team in Vietnam (27).
Of course, in addition to IDV and CCoV-HuPn-2018, public health professionals should seek to detect other animal respiratory viruses as they spill over to infect humans. When possible, such surveillance should be strategically focused at the human–animal nexus where we recognize the risk is high (31). For instance, the risk for novel swine viruses spilling over from swine to infect swine workers is exceedingly high compared with the similar risk for avian influenza viruses spilling over from poultry to infect poultry workers (32–37). Similarly, we are aware of the zoonotic threat of other animal coronaviruses infecting persons directly or indirectly exposed to their animal hosts (38,39). In addition, we posit that evidence of animal adenoviruses spilling over to infect humans is mounting (40). In recent years, molecular evidence has shown that a vampire bat–like adenovirus in Malaysia (41) and a bovine adenovirus in Pakistan (J.R.E. Ansari et al., unpub. data, https://doi.org/10.21203/rs.3.rs-5811360/v1) were associated with human respiratory disease. Because of those and other observations, we argue that periodic surveillance with targeted and panspecies diagnostics would be prudent when addressing emerging respiratory virus threats for viruses in 6 viral families (Adenoviridae, Coronaviridae, Orthomyxoviridae, Paramyxoviridae, Picornaviridae, and Pneumoviridae) (42). Conducting such surveillance in concert with occasional agnostic next-generation sequencing of specimens associated with unusual illnesses can help us better prepare for future pandemic threats at more sustainable costs than previous strategies that sought to detect novel pathogens in many wildlife hosts (43).
The novelist Stephen King adapted a classical English language quip to illustrate our tendency to ignore our previous mistakes: “Fool me once, shame on you. Fool me twice, shame on me. Fool me three times, shame on both of us” (44). As we prepare for the next pandemic, we would be wise to heed his advice.
Currently, we know of no human or veterinary laboratory-approved molecular or serologic assays for IDV or CCoV-HuPN-2018. Hence, our knowledge about the viruses’ epidemiology and clinical manifestations are limited to a modest number of research studies. Even so, the limited data regarding these novel, newly detected viruses indicate that that they are a major threat to public health. If we wish to avoid being fooled again by a novel virus suddenly gaining efficient human-to-human transmissibility and causing large human epidemics, we would be wise to develop better surveillance systems and new countermeasures for these and similar viruses. Potential actions include the development of commercial real-time reverse transcription PCR diagnostic tests specifically targeting IDV and CCoV-HuPN-2018 viruses. Conducting periodic assessments for novel respiratory viruses could detect what might be cryptic causes of hospitalizations or animal epizootics in geographic areas recognized to be sites of increased presence of emerging pathogens (42,45). As we have shown, this surveillance can easily be performed with panspecies and next-generation sequencing diagnostics (42,46,47). Clinicians should consider IDV and CCoV-HuPN-2018 in their workup of pneumonia patients when a primary battery of diagnostic tests fail to identify a pathogen. Scientists should begin evaluating antiviral drugs as effective therapy for the treatment of IDV and CCoV-HuPN-2018 infections. If further epidemiologic study indicates that the investment is warranted, human vaccine development should be considered for both IDV and CCoV-HuPN-2018. Furthermore, with respect to detecting new respiratory virus threats, when specific novel animal spillover risks are identified, they may often be mitigated with targeted interventions such as those recently reviewed by Vora et al. (48) and Plowright et al. (49).
Dr. Gray is a public health physician, infectious disease epidemiologist, and director of the One Health Research and Training Laboratory at the University of Texas Medical Branch. His research interests include emerging pathogens, especially those that are zoonotic and cause respiratory disease.
Acknowledgment
This project was supported financially by an Agriculture and Food Research Initiative competitive grant from the American Rescue Plan Act through the US Department of Agriculture’s Animal and Plant Health Inspection Service (grant no. 2023-70432-39558), the American Lung Association (grant no. Covid-862514), the US Department of Agriculture’s Agricultural Research Service (agreement no. 58-3022-4-048), and startup funding from the University of Texas Medical Branch (all awarded to G.C.G. as principal investigator).
References
- Murray CJ, Lopez AD, Chin B, Feehan D, Hill KH. Estimation of potential global pandemic influenza mortality on the basis of vital registry data from the 1918-20 pandemic: a quantitative analysis. Lancet. 2006;368:2211–8. DOIPubMedGoogle Scholar
- Viboud C, Simonsen L, Fuentes R, Flores J, Miller MA, Chowell G. Global mortality impact of the 1957–1959 influenza pandemic. J Infect Dis. 2016;213:738–45. DOIPubMedGoogle Scholar
- Honigsbaum M. Revisiting the 1957 and 1968 influenza pandemics. Lancet. 2020;395:1824–6. DOIPubMedGoogle Scholar
- Cherry JD. The chronology of the 2002-2003 SARS mini pandemic. Paediatr Respir Rev. 2004;5:262–9. DOIPubMedGoogle Scholar
- Cherry JD, Krogstad P. SARS: the first pandemic of the 21st century. Pediatr Res. 2004;56:1–5. DOIPubMedGoogle Scholar
- Dawood FS, Iuliano AD, Reed C, Meltzer MI, Shay DK, Cheng PY, et al. Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: a modelling study. Lancet Infect Dis. 2012;12:687–95. DOIPubMedGoogle Scholar
- Hause BM, Collin EA, Liu R, Huang B, Sheng Z, Lu W, et al. Characterization of a novel influenza virus in cattle and Swine: proposal for a new genus in the Orthomyxoviridae family. mBio. 2014;5:e00031–14. DOIPubMedGoogle Scholar
- Hause BM, Ducatez M, Collin EA, Ran Z, Liu R, Sheng Z, et al. Isolation of a novel swine influenza virus from Oklahoma in 2011 which is distantly related to human influenza C viruses. PLoS Pathog. 2013;9:
e1003176 . DOIPubMedGoogle Scholar - Liu R, Sheng Z, Huang C, Wang D, Li F. Influenza D virus. Curr Opin Virol. 2020;44:154–61. DOIPubMedGoogle Scholar
- Sreenivasan CC, Sheng Z, Wang D, Li F. Host range, biology, and species specificity of seven-segmented influenza viruses—a comparative review on influenza C and D. Pathogens. 2021;10:1583. DOIPubMedGoogle Scholar
- Kwasnik M, Rola J, Rozek W. Influenza D in domestic and wild animals. Viruses. 2023;15:2433. DOIPubMedGoogle Scholar
- Bailey ES, Fieldhouse JK, Alarja NA, Chen DD, Kovalik ME, Zemke JN, et al. First sequence of influenza D virus identified in poultry farm bioaerosols in Sarawak, Malaysia. Trop Dis Travel Med Vaccines. 2020;6:5. DOIPubMedGoogle Scholar
- White SK, Ma W, McDaniel CJ, Gray GC, Lednicky JA. Serologic evidence of exposure to influenza D virus among persons with occupational contact with cattle. J Clin Virol. 2016;81:31–3. DOIPubMedGoogle Scholar
- Gao H, Sun W, Lu P, Dong Z, Wu J, Li Y, et al. Efficient airborne transmission of influenza D virus in ferret models and serological evidence of human exposure in Northeast China. Emerg Microbes Infect. 2025;14:
2564308 . DOIPubMedGoogle Scholar - Limaye S, Lohar T, Dube H, Ramasamy S, Kale M, Kulkarni-Kale U, et al. Rapid evolution leads to extensive genetic diversification of cattle flu Influenza D virus. Commun Biol. 2024;7:1276. DOIPubMedGoogle Scholar
- Guerra FM Jr, Edgar EM, Oguzie JU, Marushchak LV, Pattillo M, Gray GC. Sparse evidence of influenza C on US dairy and beef cattle farms. Influenza Other Respir Viruses. 2025;19:
e70167 . DOIPubMedGoogle Scholar - Umar S, Ma MJ, Gray GC. Influenza D virus was not detected among prospectively studied swine farm in multiple provinces of China. Influenza Other Respir Viruses. 2023;17:
e13223 . DOIPubMedGoogle Scholar - Farrell AS, Bui VN, Dao TD, Hoang TD, Gray GC. No influenza D virus detected among pigs, northern Vietnam. Influenza Other Respir Viruses. 2021;15:315–7. DOIPubMedGoogle Scholar
- Pulscher LA, Maruschak LV, Shittu I, Alsharif H, Gray GC. No evidence of novel respiratory viruses on two Texas dairy farms before the H5N1 avian influenza virus epizootic. Influenza Other Respir Viruses. 2025;19:
e70146 . DOIPubMedGoogle Scholar - O’Donoghue S, Waters SM, Morris DW, Earley B. A comprehensive review: bovine respiratory disease, current insights into epidemiology, diagnostic challenges, and vaccination. Vet Sci. 2025;12:778. DOIPubMedGoogle Scholar
- Leibler JH, Abdelgadir A, Seidel J, White RF, Johnson WE, Reynolds SJ, et al. Influenza D virus exposure among US cattle workers: A call for surveillance. Zoonoses Public Health. 2023;70:166–70. DOIPubMedGoogle Scholar
- Holwerda M, Kelly J, Laloli L, Stürmer I, Portmann J, Stalder H, et al. Determining the replication kinetics and cellular tropism of influenza D virus on primary well-differentiated human airway epithelial cells. Viruses. 2019;11:377. DOIPubMedGoogle Scholar
- Vlasova AN, Diaz A, Damtie D, Xiu L, Toh TH, Lee JS, et al. Novel canine coronavirus isolated from a hospitalized patient with pneumonia in east Malaysia. Clin Infect Dis. 2022;74:446–54. DOIPubMedGoogle Scholar
- Lednicky JA, Tagliamonte MS, White SK, Blohm GM, Alam MM, Iovine NM, et al. Isolation of a novel recombinant canine coronavirus from a visitor to Haiti: further evidence of transmission of coronaviruses of zoonotic origin to humans. Clin Infect Dis. 2022;75:e1184–7. DOIPubMedGoogle Scholar
- Theamboonlers A, Samransamruajkit R, Thongme C, Amonsin A, Chongsrisawat V, Poovorawan Y. Human coronavirus infection among children with acute lower respiratory tract infection in Thailand. Intervirology. 2007;50:71–7. DOIPubMedGoogle Scholar
- Silva CS, Mullis LB, Pereira O, Saif LJ, Vlasova A, Zhang X, et al. Human respiratory coronaviruses detected in patients with influenza-like illness in Arkansas, USA. Virol Mycol. 2014;2014(Suppl 2). PMID 27588218
- Phan PT, Wijayaratne G, Bodinayake CK, Oguzie JU, Nguyen-Tien T, Marushchak LV, et al. Diverse respiratory viruses detected among hospitalized patients with pneumonia in Sri Lanka and Vietnam. IJID Reg. 2025;17:
100757 . DOIPubMedGoogle Scholar - Tortorici MA, Walls AC, Joshi A, Park YJ, Eguia RT, Miranda MC, et al. Structure, receptor recognition, and antigenicity of the human coronavirus CCoV-HuPn-2018 spike glycoprotein. Cell. 2022;185:2279–2291.e17. DOIPubMedGoogle Scholar
- Liu Y, Chen D, Wang Y, Li X, Qiu Y, Zheng M, et al. Characterization of CCoV-HuPn-2018 spike protein-mediated viral entry. J Virol. 2023;97:
e0060123 . DOIPubMedGoogle Scholar - Wang F, Yang G, Yan L. Crystal structures of fusion cores from CCoV-HuPn-2018 and SADS-CoV. Viruses. 2024;16:272. DOIPubMedGoogle Scholar
- Borkenhagen LK, Wang GL, Simmons RA, Bi ZQ, Lu B, Wang XJ, et al. High risk of influenza virus infection among swine workers: examining a dynamic cohort in China. Clin Infect Dis. 2020;71:622–9. DOIPubMedGoogle Scholar
- Ma M, Anderson BD, Wang T, Chen Y, Zhang D, Gray GC, et al. Serological evidence and risk factors for swine influenza infections among Chinese swine workers in Guangdong Province. PLoS One. 2015;10:
e0128479 . DOIPubMedGoogle Scholar - Myers KP, Olsen CW, Setterquist SF, Capuano AW, Donham KJ, Thacker EL, et al. Are swine workers in the United States at increased risk of infection with zoonotic influenza virus? Clin Infect Dis. 2006;42:14–20. DOIPubMedGoogle Scholar
- Khuntirat BP, Yoon IK, Blair PJ, Krueger WS, Chittaganpitch M, Putnam SD, et al. Evidence for subclinical avian influenza virus infections among rural Thai villagers. Clin Infect Dis. 2011;53:e107–16. DOIPubMedGoogle Scholar
- Kayali G, Ortiz EJ, Chorazy ML, Gray GC. Evidence of previous avian influenza infection among US turkey workers. Zoonoses Public Health. 2010;57:265–72. DOIPubMedGoogle Scholar
- Borkenhagen LK, Salman MD, Ma MJ, Gray GC. Animal influenza virus infections in humans: A commentary. Int J Infect Dis. 2019;88:113–9. DOIPubMedGoogle Scholar
- Sánchez CA, Li H, Phelps KL, Zambrana-Torrelio C, Wang LF, Zhou P, et al. A strategy to assess spillover risk of bat SARS-related coronaviruses in Southeast Asia. Nat Commun. 2022;13:4380. DOIPubMedGoogle Scholar
- Vlasova AN, Toh TH, Lee JS, Poovorawan Y, Davis P, Azevedo MSP, et al. Animal alphacoronaviruses found in human patients with acute respiratory illness in different countries. Emerg Microbes Infect. 2022;11:699–702. DOIPubMedGoogle Scholar
- Borkenhagen LK, Fieldhouse JK, Seto D, Gray GC. Are adenoviruses zoonotic? A systematic review of the evidence. Emerg Microbes Infect. 2019;8:1679–87. DOIPubMedGoogle Scholar
- Fieldhouse JK, Bailey ES, Toh TH, Hii KC, Mallinson KA, Ting J, et al. Panspecies molecular assays detect viral pathogens missed by real-time PCR/reverse-transcriptase PCR among pneumonia patients, Sarawak, Malaysia. Trop Dis Travel Med Vaccines. 2020;6:13. DOIPubMedGoogle Scholar
- Gray GC, Robie ER, Studstill CJ, Nunn CL. Mitigating future respiratory virus pandemics: new threats and approaches to consider. Viruses. 2021;13:637. DOIPubMedGoogle Scholar
- Aatresh AV, Lipsitch M. What is the relationship between viral prospecting in animals and medical countermeasure development? mBio. 2025;16:
e0203325 . DOIPubMedGoogle Scholar - King S. On writing: a memoir of the craft. New York: Scribner; 2000.
- Gray GC, Abdelgadir A. While we endure this pandemic, what new respiratory virus threats are we missing? Open Forum Infect Dis. 2021;8:ofab078. DOIGoogle Scholar
- Shittu I, Oguzie JU, Hernández-Vidal G, Moreno-Degollado G, Silva DB, Marushchak LV, et al. Novel rodent coronavirus-like virus detected among beef cattle with respiratory disease in Mexico. Viruses. 2025;17:433. DOIPubMedGoogle Scholar
- Oguzie JU, Hernandez-Vidal G, Moreno-Degollado G, Gray GC. First detection of boosepivirus B1 in a sick yearling’s nasal swab, Mexico. Influenza Other Respir Viruses. 2025;19:
e70165 . DOIPubMedGoogle Scholar - Vora NM, Hannah L, Walzer C, Vale MM, Lieberman S, Emerson A, et al. Interventions to reduce risk for pathogen spillover and early disease spread to prevent outbreaks, epidemics, and pandemics. Emerg Infect Dis. 2023;29:1–9. DOIPubMedGoogle Scholar
- Plowright RK, Ahmed AN, Coulson T, Crowther TW, Ejotre I, Faust CL, et al. Ecological countermeasures to prevent pathogen spillover and subsequent pandemics. Nat Commun. 2024;15:2577. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: January 21, 2026
Table of Contents – Volume 32, Number 1—January 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Gregory C. Gray, University of Texas Medical Branch, 301 University Blvd, Rte 0435, Galveston, TX 77555-0435, USA
Top