Volume 32, Number 7—July 2026
Dispatch
Phormia regina Fly as Vector for Ignatzschineria spp. Bacteremia in Persons Experiencing Homelessness, Canada, 2025
Cite This Article
Citation for Media
Abstract
Ignatzschineria spp. bacteria are emerging pathogens whose vectors historically have not been clearly identified. We used molecular methods to establish a relationship between the black blow fly (Phormia regina) and human Ignatzschineria bacteremia in persons experiencing homelessness in Vancouver, British Columbia, Canada, validating a novel transmission pathway in a vulnerable urban population.
Ignatzschineria bacteria are increasingly recognized as a cause of human infection (1). Four species have been described to date: I. indica, I. larvae, I. ureiclastica, and I. cameli (1). Historically, species associated with human wound and blood infection have shown a geographic distribution; I. indica predominates in North America, and I. larvae/I. ureiclastica predominates in Europe (1). The gram-negative, aerobic, nonmotile, and oxidase- and catalase-positive bacteria were originally isolated from the larvae and adult gastrointestinal tracts of Wohlfahrtia magnifica parasitic flies (2). Experimental and ecologic studies suggest that Ignatzschineria are part of the larvae microbiome through a combination of vertical transmission and environmental acquisition during feeding (3–5). Ultimately, host identity appears to shape overall abundance; blow fly species such as Phormia regina have a high abundance of Ignatzschineria (Appendix Figure 1) (4).
Although W. magnifica flies primarily cause myiasis in animals, blow flies are associated with facultative myiasis in humans (6). At least 3 case reports of Ignatzschineria bacteremia in humans have implicated the green blow fly (Lucilia sericata), but the methods used for identification have primarily relied on epidemiology and morphologic features (1,7). In general, the specific fly vectors associated with human urban myiasis are seldom confirmed molecularly.
We describe 2 cases of Ignatzschineria bacteremia in persons experiencing homelessness (PEH) in Vancouver, British Columbia, Canada. We used genomic approaches to characterize the bacterial isolates from both cases and performed vector identification on the fly associated with 1 of the cases. We obtained written consent from both patients for publication of their cases.
Figure 1
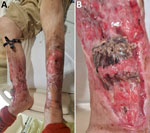
Figure 1. Leg wounds in case-patient 1 in pair of Ignatzschineriaspp. bacteremia cases in persons experiencing homelessness, Vancouver, British Columbia, Canada, 2025. A) Bilateral leg wounds at time of admission;...
Case 1 involved a 49-year-old man, a PEH with active substance use disorder (SUD) and chronic bilateral leg wounds, who sought care for worsening pain, swelling, and wound myiasis (Figure 1, panel A, B). We collected 2 aerobic and 2 anaerobic blood cultures, and within 24 hours, all 4 collection bottles grew gram-negative bacilli. After an additional 24 hours, small gray colonies appeared on sheep blood agar and clear colonies appeared on MacConkey agar (Appendix Figure 2). Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI Biotyper; Bruker, https://www.bruker.com) identified the isolates as I. larvae, I. indica, and Fusobacterium varium. Whole-genome sequencing (WGS) confirmed the species as I. larvae and I. indica (Appendix Figure 3).
We conducted antimicrobial susceptibility testing by using gradient strip diffusion method for several antibiotics (Table). We referenced the 2025 Clinical and Laboratory Standards Institute breakpoints for other non-Enterobacterales for interpretation of MICs (8). The genotypic antimicrobial resistance prediction for the I. larvae isolate was concordant with phenotype for all tested antimicrobials (Table). The I. indica isolate did not grow for phenotypic testing, but no genotypic resistance was detected for antibiotics of interest.
The patient underwent surgical debridement of both leg wounds and a split thickness skin grafting from the upper thigh. Computed tomography imaging of the right lower leg demonstrated periosteal reaction and irregularity of the underlying bone cortex. He was treated for osteomyelitis with 6 weeks of amoxicillin/clavulanate. After antimicrobial therapy, the leg wounds healed well and showed healthy granulation tissue. We performed no repeat imaging.
Figure 2
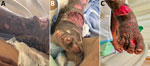
Figure 2. Leg wounds in case-patient 2 in pair of Ignatzschineriaspp. bacteremia cases in persons experiencing homelessness, Vancouver, British Columbia, Canada, 2025. A, B) Lateral (A) and anterior (B) right...
Shortly after case-patient 1 was admitted to hospital, a second patient (case-patient 2), a 36-year-old man who was a PEH and SUD, sought care for fevers and an erythematous right shin containing necrotic wounds heavily infested with fly larvae (Figure 2, panel A, B). His blood cultures grew I. larvae and Pasteurella multocida, which we confirmed by using WGS. The genotypic antimicrobial resistance prediction was concordant with phenotype for all antibiotics tested (Table). His orthopedic and plastic surgery physicians recommended wound care without surgical intervention. The plan was to complete 6 weeks of ceftriaxone for osteomyelitis, but the patient self-initiated discharge before completing therapy.
Figure 3
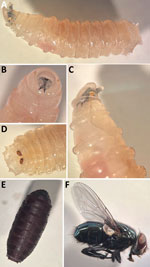
Figure 3. Development of black blow fly (Phormia regina) from larvae collected from case-patient 2 in pair of Ignatzschineriaspp. bacteremia cases in persons experiencing homelessness, Vancouver, British Columbia,...
When case-patient 2 sought care, we recognized a pattern of myiasis-associated bacteremia. We collected fly larvae from the patient in a sterile container, stored them on blood agar, and observed them over time to document their development into adult flies (Figure 3). The third instar larvae (9–12 mm) were creamy white with distinct bands along the body covered in short spines. The prothoracic spiracles had >10 openings. We observed 2 incomplete peritremes on the posterior with three inner slits directed toward the median line ventrally. The adult flies (10–12 mm) were metallic green. Considered together, those features were suggestive of the Calliphoridae family and the Phormia genus (9). We confirmed the fly to be P. regina by sequencing the cytochrome c oxidase subunit I gene using universal primers (Appendix Figure 4) (10). To confirm the presence of Ignatzschineria spp. in the fly larvae, we washed a larva 3 times in phosphate-buffered saline, macerated the anterior portion of the larva in phosphate-buffered saline, and subjected the homogenate to mechanical disruption with glass beads before DNA extraction and 16S rRNA sequencing. We uploaded sequencing data to the CZ ID platform (https://czid.org) for metagenomic analysis (S.E. Simmonds et al., unpub. data, https://doi.org/10.1101/2024.02.29.579666). We identified Ignatzschineria spp. at a 16S rRNA read abundance of 0.1% from the larva bacterial microbiome, providing molecular evidence of vector colonization.
We have established a relationship between P. regina fly wound myiasis and I. larvae bacteremia by identifying P. regina flies as the probable causative agent of myiasis, isolating Ignatzschineria bacteria in patients’ blood cultures, and detecting the organism within the larval microbiome. Our findings fulfill the criteria for sequence-based determination of causation as proposed by Fredricks and Relman (11). Moreover, the larval microbiome results were consistent with the findings of Deguenon et al. (12), who identified Ignatzschineria bacteria in the microbiome of wild P. regina flies. Because P. regina blow flies are ubiquitous and widely distributed across North America, our results suggest that exposure risk could be far broader than previously assumed.
Both patients were at heightened risk for Ignatzschineria bacteremia because of their PEH status, SUD, chronic wounds, and competing health and social priorities (i.e., resuming everyday life versus receiving treatment and follow-up) (13). We hypothesize that the complexity of wounds and the magnitude of infestation might increase the risk for Ignatzschineria infection. Those cases also illustrate the value of a One Health perspective, in which human vulnerability, environmental exposure, and interactions with urban flies converge to create conditions for pathogen transmission. Clinicians should maintain a high index of suspicion and obtain blood cultures in patients with wound myiasis and systemic symptoms.
Accurate identification of Ignatzschineria bacteria remains a diagnostic challenge in the clinical microbiology laboratory. In several cases, Ignatzschineria isolates were initially misidentified as other bacteria using phenotypic and molecular methods (7,13). We observed that the Bruker MALDI Biotyper successfully identified I. indica and I. larvae/ureiclastica but could not differentiate between I. larvae and I. ureiclastica because of the high degree of genetic similarity. Of note, the VITEK MS version 3.2 knowledge base (bioMérieux, https://www.biomerieux.com) does not include Ignatzschineria species. Most case reports have relied on molecular methods such as 16S rRNA gene sequencing for genus identification, but only WGS can truly provide species-level resolution (14). Those findings highlight a broader diagnostic gap in detecting emerging zoonotic pathogens and demonstrate the value of WGS for achieving species-level resolution.
Information on the antimicrobial susceptibility of Ignatzschineria is currently limited. Reported testing methods include disk diffusion, automated instruments, gradient strip diffusion, or some combination of those methods (1). Most published cases demonstrate that Ignatzschineria bacteria are susceptible to β-lactam antibiotics, which comprise the mainstay of therapy (1). Treatment typically includes appropriate antimicrobial therapy combined with larvae removal, wound care, debridement, amputation, or some combination of these interventions (1,7). Overall, patient outcomes have been favorable when both infection and underlying wounds are addressed (13).
Dr. Finlayson-Trick is a third-year medical microbiology resident at the University of British Columbia, Vancouver. Her primary research interests include antimicrobial resistance with a focus on detection in the clinical microbiology laboratory, optimization of antimicrobial stewardship strategies, and advancement of novel therapeutics.
Acknowledgments
We thank Cherie Lai, Gillian Andersen, and the rest of the microbiology laboratory staff at St. Paul’s Hospital who played an instrumental role in rearing the fly larvae.
Sequence data from this study are available in GenBank (BioProject accession no. PRJNA1438763).
Author contributions: review and editing, E.C.L.F.-T., A.F.A., Y.Z.J., M.P., G.R., A.S., C.F.L., S.D.C., N.M., P.T., V.L., D.H., and M.G.R.; supervision, M.G.R.; original draft, E.C.L.F.-T., A.F.A., Y.Z.J., M.P., A.S., C.F.L., and N.M.; methodology, E.C.L.F.-T., M.P., G.R., and P.T.; project administration, M.P., P.T., and M.G.R.; investigation, E.C.L.F.-T., M.P., G.R., A.S., C.F.L., S.D.C., N.M., P.T., and M.G.R.; formal analysis, E.C.L.F.-T., M.P., G.R., S.D.C., P.T., and M.G.R.; data curation, E.C.L.F.-T., M.P., G.R., S.D.C., and P.T.; conceptualization, E.C.L.F.-T., A.F.A., Y.Z.J., M.P., G.R., A.S., C.F.L., N.M., P.T., V.L., D.H., and M.G.R.
References
- Fear T, Richert Q, Levesque J, Walkty A, Keynan Y. Ignatzschineria indica bloodstream infection associated with maggot infestation of a wound in a patient from Canada. J Assoc Med Microbiol Infect Dis Can. 2020;5:193–200. DOIPubMedGoogle Scholar
- Tóth E, Kovács G, Schumann P, Kovács AL, Steiner U, Halbritter A, et al. Schineria larvae gen. nov., sp. nov., isolated from the 1st and 2nd larval stages of Wohlfahrtia magnifica (Diptera: Sarcophagidae). Int J Syst Evol Microbiol. 2001;51:401–7. DOIPubMedGoogle Scholar
- Long W, Pang J, Yan W, Hu N. Industrial-scale bioconversion of three-phase residue by Musca domestica larvae: dynamics of gut microbiota and their ecological driver. Insects. 2025;16:686. DOIPubMedGoogle Scholar
- Mikaelyan A, Receveur J, Bernstein K, Babcock NJ, Pechal JL, Welsh MV, et al. Host-specific microbiomes of blow flies: ecological drivers and implications for pathogen carriage. Front Immunol. 2025;16:
1673934 . DOIPubMedGoogle Scholar - Tang H, Zhang X, Yang F, Zhang C, Ngando FJ, Ren L, et al. Effect of ciprofloxacin on the composition of intestinal microbiota in Sarcophaga peregrina (Diptera: Sarcophagidae). Microorganisms. 2023;11:2867. DOIPubMedGoogle Scholar
- Cardoso GA, Cunha VAS, Genevcius BC, Madeira-Ott T, Costa BMA, Rossoni DM, et al. Origins and diversification of myiasis across blowflies. Ecol Evol. 2025;15:
e70993 . DOIPubMedGoogle Scholar - Barker HS, Snyder JW, Hicks AB, Yanoviak SP, Southern P, Dhakal BK, et al. First case reports of Ignatzschineria (Schineria) indica associated with myiasis. J Clin Microbiol. 2014;52:4432–4. DOIPubMedGoogle Scholar
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing, 35th edition. Supplement M100. Wayne (PA): The Institute; 2025.
- Centers for Disease Control and Prevention. Pictorial keys: arthropods, reptiles, birds, and mammals of public health significance. 2000 May 22 [cited 2025 Sep 23]. https://stacks.cdc.gov/view/cdc/60114
- Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.PubMedGoogle Scholar
- Fredricks DN, Relman DA. Sequence-based identification of microbial pathogens: a reconsideration of Koch’s postulates. Clin Microbiol Rev. 1996;9:18–33. DOIPubMedGoogle Scholar
- Deguenon JM, Travanty N, Zhu J, Carr A, Denning S, Reiskind MH, et al. Exogenous and endogenous microbiomes of wild-caught Phormia regina (Diptera: Calliphoridae) flies from a suburban farm by 16S rRNA gene sequencing. Sci Rep. 2019;9:20365. DOIPubMedGoogle Scholar
- Pomerantz E, Pericak O, Sokach C, Edathil J, Yeung HM. Two cases of maggot‐associated Ignatzschineria bacteremia in xylazine‐induced injection wounds: an emerging threat. Case Rep Infect Dis. 2024;2024:
7684187 . DOIPubMedGoogle Scholar - Belote A, Hawkinson D, Shoemaker DM. Myiasis as a vector for bacteremia: a unique case of Helcococcus kunzii and Ignatzschineria ureiclastica/larvae polymicrobial bacteremia from myiasis. Vector Borne Zoonotic Dis. 2024;24:788–91. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: May 19, 2026
Table of Contents – Volume 32, Number 7—July 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Emma Finlayson-Trick, University of British Columbia Medical Microbiology Residency Program, Department of Pathology and Laboratory Medicine, University of British Columbia, 910 West 10th Ave, Vancouver, BC V5Z 1M9, Canada
Top