Volume 9, Number 11—November 2003
Research
Triosephosphate Isomerase Gene Characterization and Potential Zoonotic Transmission of Giardia duodenalis
Cite This Article
Citation for Media
Abstract
To address the source of infection in humans and public health importance of Giardia duodenalis parasites from animals, nucleotide sequences of the triosephosphate isomerase (TPI) gene were generated for 37 human isolates, 15 dog isolates, 8 muskrat isolates, 7 isolates each from cattle and beavers, and 1 isolate each from a rat and a rabbit. Distinct genotypes were found in humans, cattle, beavers, dogs, muskrats, and rats. TPI and small subunit ribosomal RNA (SSU rRNA) gene sequences of G. microti from muskrats were also generated and analyzed. Phylogenetic analysis on the TPI sequences confirmed the formation of distinct groups. Nevertheless, a major group (assemblage B) contained most of the human and muskrat isolates, all beaver isolates, and the rabbit isolate. These data confirm that G. duodenalis from certain animals can potentially infect humans and should be useful in the detection, differentiation, and taxonomy of Giardia spp.
Giardiasis is a common cause of diarrheal disease in almost all vertebrates, including humans. In industrialized countries, it is referred to as a reemerging infectious disease because of its increasingly recognized role in outbreaks of diarrheal disease in daycare centers and in water- and foodborne outbreaks. Giardia is also one of the most frequently observed parasites infecting dairy cattle and domestic dogs. In developing countries in Asia, Africa, and Latin America, approximately 200 million people have symptomatic giardiasis (1).
The taxonomy of Giardia at the species level is complicated and unresolved because of limited morphologic differences. Based on morphology, six species of this genus are considered valid: Giardia duodenlalis (syn. G. lamblia or G. intestinalis) in a wide range of mammals, including humans, livestock, and companion animals; G. agilis in amphibians; G. muris in rodents; G. ardeae and G. psittaci in birds; and G. microti in muskrats and voles (2–6). However, on the basis of host origins, 41 Giardia species have been named (7,8).
Molecular tools have been used recently to characterize the epidemiology of human giardiasis. Although isolates of G. duodenalis from humans and various animals are morphologically similar, distinct host-adapted genotypes have been demonstrated within G. duodenalis (1,9–12). Two major groups of G. duodenalis have been recognized as infecting humans worldwide, but there are some differences in naming of these groups, as evidenced by the following categorizations: Polish and Belgian genotypes (9); groups 1, 2, and 3 (10,13); and assemblages A and B (11). So far, no general consensus has been reached concerning the nomenclature of these genotypes, but the term assemblages has been more widely used. The finding of host-adapted Giardia genotypes is of public health importance, considering the controversy regarding the zoonotic potential of Giardia (1,14).
We describe the development of a two-step nested polymerase chain reaction (PCR) protocol to amplify the triosephosphate isomerase (TPI) gene of G. duodenalis and G. microti and nucleotide sequence characterization of amplified TPI fragment. The TPI gene was chosen because of the high genetic heterogeneity displayed by Giardia spp. at this locus (12,15). Results of the study have validated previous observations on the genetic diversity of Giardia parasites on the basis of characterization of the glutamate dehydrogenase (GDH), small subunit ribosomal RNA (SSU rRNA), and TPI genes (12,15–17). These data also suggest that some animal isolates of G. duodenalis are of zoonotic potential. These data should be useful in developing alternative molecular tools to differentiate Giardia parasites at species and genotype levels and in investigating giardiasis outbreaks or endemic diseases.
G. duodenalis Isolates and DNA Extraction
Fecal samples containing G. duodenalis cysts were obtained from infected humans, cattle, companion animals (dogs and a rabbit), aquatic wildlife (beavers and muskrats), and one rat. Human samples were mostly from sporadic cases, with the exception of two isolates (4599, 4600) from a foodborne outbreak. Fecal samples with G. microti were obtained from infected muskrats. Giardia infection was diagnosed by microscopy of wet mounts or immunofluorescence-stained materials. Samples were stored at 4°C in 2.5% (w/v) potassium dichromate solution or frozen at –20°C and used in DNA extraction without cyst isolation (Table 1). For DNA extraction, 200 μL of the fecal suspension from each sample was aliquoted and washed three times with distilled water. The material was treated initially with 66.7 μL of 1 M KOH and 18.6 μL of 1 M dithiothreitol (DTT) followed by neutralization with 8.6 μL of 25% (v/v) hydrochloric acid. The DNA lysate was then extracted once with phenol:chloroform:isoamyl alcohol (25:24:1) solution, and purified by using the QIAamp DNA Stool Kit (QIAGEN Inc, Valencia, CA). Because the extracted DNA contained nucleic acids from both Giardia cysts and fecal materials, DNA concentration was not determined for all samples.
PCR Amplification of the TPI Gene
To amplify the TPI fragment from various Giardia isolates, a nested PCR protocol was developed that used primers complementary to the conserved published TPI nucleotide sequences of various Giardia parasites downloaded from GenBank: G. duodenalis (U57897, AF06957 to AF069563, L02116, L02120), G. muris (AF069565), and G. ardeae (AF069564). For the primary PCR, a PCR product of 605 bp was amplified by using primers AL3543 [5′-AAATIATGCCTGCTCGTCG-3′] and AL3546 [5′-CAAACCTTITCCGCAAACC-3′]. The PCR reaction comprised 0.25–2.0 μL of DNA, 200 μM each of deoxynucleoside triphosphate (dNTP), 1X PCR buffer (Perkin Elmer, Wellesley, MA), 3.0 mM MgCl2, 5.0 U of Taq polymerase (GIBCO BRL, Frederick, MD), and 200 nM of each primer in a total of 100-μL reaction. The reactions were performed for 35 cycles (94°C for 45 s, 50°C for 45 s, and 72°C for 60 s) in a Perkin-Elmer GeneAmp PCR 9700 thermocycler, with an initial hot start (94°C for 5 min) and a final extension (72°C for 10 min). For the secondary PCR, a fragment of 530 bp was amplified by using 2.5 μL of primary PCR reaction and primers AL3544 [5′-CCCTTCATCGGIGGTAACTT-3′] and AL3545 [5′-GTGGCCACCACICCCGTGCC-3′]. The conditions for the secondary PCR were identical to the primary PCR. The PCR products were analyzed by agarose gel electrophoresis and visualized after ethidium bromide staining.
PCR Amplification of the SSU rRNA Gene
A nested PCR protocol was also developed to amplify the SSU rRNA fragment from Giardia isolates, using primers complementary to the conserved published SSU rRNA nucleotide sequences from various Giardia parasites downloaded from GenBank: G. duodenalis (AJ278959, AJ293295 to AJ293299, AJ293300, AJ293301, L29129, M54878, U09491, U09492, X52949), G. microti (AF006676, AF006677), G. muris (X65063), and G. ardeae (Z17210). For the primary PCR, a PCR product of 300 bp was amplified by using primers AL4303 [5′-ATCCGGTCGATCCTGCCG-3′] and reverse AL4305 [5′-AGGATCAGGGTTCGACT-3′]. The PCR reaction was performed by using the GC-RICH PCR System kit, which consisted of GC-RICH Enzyme mix (Taq polymerase in combination with a proofreading polymerase), GC-RICH-PCR reaction buffer (includes a final 1.5 mM MgCl2 and dimethyl sulfoxide [DMSO]), and GC-RICH resolution solution (Roche Diagnostics, Indianapolis, IN) with 0.25–2.0 μL of DNA, 200 μM each of dNTP, and 200 nM of each primer in a total of 50-μL reaction. For the secondary PCR, a fragment of 255 bp was amplified with the GC-RICH PCR System kit (Roche) with 2.5 μL of primary PCR reaction, and 200 nM of primers AL4304 [5′-CGGTCGATCCTGCCGGA-3′] and AL4306 [5′-GGCGGAGGATCAGGGT-3′]. The cycling conditions for both SSU RNA primary and secondary PCR were identical to those used to amplify the TPI gene.
DNA Sequencing and Phylogenetic Analysis
The secondary PCR products were purified by using Microcon PCR Centrifugal Filter Devices (Millipore Corp., Bedford, MA) and sequenced on an ABI 3100 automated sequencer by using the Big Dye Terminator Cycle Sequencing Ready Reaction Kit (Perkin-Elmer). Sequence accuracy was confirmed by two-directional sequencing of two separate PCR products. Multiple alignment of the nucleotide sequences was performed by using Wisconsin Package Version 9.0 program (18). A phylogenetic analysis was performed on the aligned sequences to assess the extent of genetic diversity within G. duodenalis parasites as well as their evolutionary relationships with other Giardia species. In this analysis, published TPI nucleotide sequences representing G. duodenalis (from humans, cattle, cat, dog, muskrat, pig, and rat), G. muris, and G. ardeae were aligned with TPI sequences of Giardia parasites obtained in this study.
A neighbor-joining tree (19) was constructed on the basis of the evolutionary distances calculated by the Kimura-2-parameter model using the TreeconW program (20). A sequence of G. ardeae (GenBank accession no. AF069564) was used as the outgroup since the construction of an unrooted tree showed it to be the most divergent member under analysis. The reliability of these trees was assessed by using the bootstrap method (21) with 1,000 pseudoreplicates; values >70% were reported (22). Nucleotide sequences of the TPI gene of G. duodenalis from humans, cattle, dogs, muskrat, rat, and rabbit, representing different genotypes, were deposited in GenBank under accession numbers AY228628 to AY228649.
A similar phylogenetic analysis was carried out on the nucleotide sequences of the SSU rRNA gene from G. microti in muskrats. SSU rRNA nucleotide sequences were deposited in GenBank under accession numbers AY228332 and AY228333.
PCR products of the expected size (approximately 500 bp) were generated from all 76 isolates. All were sequenced, and all of the nucleotide sequences obtained belonged to the TPI sequences of Giardia based on BLAST search of the GenBank database. The sources of these isolates were humans (37 isolates), dogs (15 isolates), muskrats (8 isolates), cattle (7 isolates), beavers (7 isolates), rabbit (1 isolate) and rat (1 isolate). The TPI gene of Giardia parasites was rich in GC content, ranging from 50.1% to 58.2% (Table 1). Isolates within each genotype, however, had very similar GC contents in the TPI gene.
The extent of genetic diversity in the genus Giardia was assessed by multiple alignments of the TPI nucleotide sequences followed by estimates of genetic distances (Table 2). The analysis showed distinct sequences for the human, cattle, beaver, dog, muskrat, and rat isolates; most animals had one genotype, and humans and muskrats had two genotypes. The genetic polymorphism in Giardia parasites was evident along the entire TPI gene both at the interspecies (Giardia spp.) and intraspecies (G. duodenalis) levels.
Figure 1
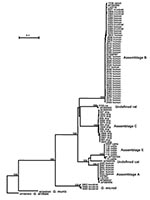
Figure 1. Phylogenetic relationships of Giardia parasites inferred by the neighbor-joining analysis of the triosephosphate isomerase (TPI) nucleotide sequences.
To understand the genetic structure of Giardia parasites, a neighbor-joining tree was constructed in a phylogenetic analysis of aligned TPI gene sequences of various Giardia species and G. duodenalis genotypes; we used the nucleotide sequence of G. ardeae (AF069564) as an outgroup to root the tree (Figure 1). The phylogenetic analysis showed four distinct clusters for the genus Giardia. The first cluster consisted of all isolates of G. duodenalis from various sources (humans, cattle, cats, dogs, beavers, muskrats, pigs, and rats). The second cluster consisted of some of the isolates from muskrats. The third and fourth cluster was each represented by a single published sequence of G. muris (AF069565) and G. ardeae (AF069564).
Several large groups were in the G. duodenalis cluster. A major group (assemblage B) was formed with most of the human and muskrat isolates, all the isolates from beavers, and the rabbit isolate (Figure 1). The remaining human isolates aligned with other previously reported human TPI sequences and formed a distinct cluster (assemblage A). Distinct clusters were also evident for the isolates from dogs (assemblage C) and rats (undefined). The cattle sequences, together with the published pig TPI sequence, also formed a distinct cluster (assemblage E or hoofed livestock genotype). Phylogenetic analysis indicated that assemblages B and C and the rat genotype were related to each other and that assemblages A and E and the cat genotype were related to each other. The formation of all major groups was supported by bootstrap analysis with full statistical reliability (Figure 1).
Figure 2

Figure 2. Variation in the triosephosphate isomerase (TPI) nucleotide sequences of G. duodenalis isolates belonging to the assemblage B. Twelve distinct subtypes of G. duodenalis based on the these sequences were evident within...
Intragenotypic variations were evident within assemblages A, B, C, and E (Table 3). A very high degree of polymorphism was noticed within the isolates from humans. The human isolates grouped in assemblage B had five SNPs (single nucleotide polymorphisms): A or G at position 39, C or T at position 91, G or A at position 162, C or T at position 165, and C or T at position 168 (position numbers according to the GenBank accession no. L02116). Within assemblage B, 12 subtypes of G. duodenalis were noticed; 11 of these had not been reported before (Figure 2). No genetic polymorphism was evident in the TPI sequences of the beaver isolates characterized so far, which were identical to those from most muskrats belonging to the major assemblage B group. However, two muskrat isolates (3565, 3569) in assemblage B had one SNP at position 216 (C to T). Six SNPs (A or G at position 51, T or C at position 77, T or G at position 150, C or T at position 330, T or C at position 383, and C or A at position 393) were evident within the dog isolates (assemblage C). Multiple alignments of sequences from hoofed livestock showed two distinct subtypes in cattle with four SNPs (T or C at position 72, G or T at position 78, T or C at position 93, and G or A at position 109). The sequence from the rat matched with the TPI sequence from another suckling mouse (GenBank accession no. AF069562) with one SNP (G to A) at position 54. No genetic variation was observed in the human TPI sequences of assemblage A generated in this study, even though three sequences from GenBank (AF069556, L02120, and U57897) had three SNPs.
Figure 3

Figure 3. Genetic variation in the nucleotide sequences of Giardia microti parasites in the small subunit ribosomal RNA (SSU rRNA) (a) and triosephosphate isomerase (TPI) (b) genes.
Since TPI nucleotide sequences of three isolates from muskrats were very different from known G. duodenalis isolates or with C. muris or C. ardeae and since they formed a distinct cluster, these isolates were characterized at the SSU rRNA locus. The Giardia SSU rRNA sequences obtained were aligned with the published sequences. Analysis showed that these isolates were G. microti. Of the three muskrat SSU rRNA sequences from this study, two (isolates 3460 and 3464) were identical to a published SSU rRNA sequence (AF006676) from muskrats (6). The third sequence (isolate 3463) was unique and had three SNPs compared with the other two muskrat isolates and AF006676. Isolate 3463 was still considered to be a sequence of G. microti because another published G. microti sequence (AF006677) was even more divergent (Figure 3a). A similar pattern of genetic polymorphism was evident in the TPI gene; the sequences from isolates 3460 and 3464 were identical to each other but had six SNPs compared with isolate 3463. This finding suggests that at least two distinct genotypes of G. microti were present in muskrats (Figure 3b).
Understanding the taxonomic relationship of a particular group of protozoan parasites that truly reflects biological characteristics and evolutionary relationships is difficult. Most protozoan parasites lack fossil records, are microscopic, and have few informative morphologic and ultrastructural characters; some lack sexual reproduction (23,24). Although Giardia spp. populate the intestinal tracts of almost every group of vertebrates, G. duodenalis is the only species found in humans and many other mammals including cattle, cats, dogs, horses, sheep, and pigs (1,25,26). Giardia cysts have also been detected in various wild mammals (14,27–34). Although these wild mammals are generally assumed to be infected with G. duodenalis, molecular characterization to support this supposition is lacking.
Even though Giardia isolates from different mammalian hosts were similar in form, a marked biological diversity among these isolates was noticed in host infectivity (35), metabolism (36), and in vitro and in vivo growth requirements (37,38). Multilocus enzyme electrophoresis identified a number of distinct groups of G. duodenalis (39,40). The forgoing heterogeneity suggests that G. duodenalis is a species-complex (39,41,42). Phylogenetic characterization based on the nucleotide sequences of GDH, elongation factor 1α (EF1α), TPI, and SSU rRNA genes suggests the presence of five to seven lineages of G. duodenalis (12,17,43,44). Among the loci analyzed, TPI has the highest degree of polymorphism (12). However, only four isolates from humans, two isolates from mice, and one isolate each from cat, dog, pig, rat (G. muris), and blue heron (G. ardeae) have been characterized at the TPI locus (12).
The genetic relationship among various Giardia parasites showed by phylogenetic analysis of the TPI gene in this study is largely in agreement with previous observations based on results from the SSU rRNA, TPI, GDH, and EF1α genes (12,17,43,44). Thus, on the basis of published and present TPI nucleotide sequences, the following groupings of G. duodenalis parasites are evident by all analyses with strong statistical reliability: 1) the formation of a group containing relatively few human isolates (assemblage A); 2) a major group containing most of the human and muskrats isolates, as well as isolates from beavers and a rabbit (assemblage B); 3) the formation of a group containing all isolates from cattle and pigs (assemblage E or the hoofed livestock genotype); 4) the formation of a group containing isolates from dogs (assemblage C); 5) an undefined cat genotype; and 6) an undefined genotype from rats. The assemblage D previously seen in a few dogs (42) was not found in this study.
In our study, a distinct and more distant cluster was formed by some isolates from muskrats. DNA sequence analysis of the SSU rRNA gene indicated that these isolates were G. microti. This organism was placed between the clades representing the G. muris and all the six assemblages of G. duodenalis. Giardia microti was established as a separate species because of sequence uniqueness of the SSU rRNA gene and minor morphologic differences from G. duodenalis (5,6). Our characterization of TPI nucleotide sequences from muskrats supports the validity of G. microti.
Results of phylogenetic analysis are useful in understanding the public health importance of some G. duodenalis parasites. Human G. duodenalis are placed in two distinct lineages (assemblages A and B), whereas the other four lineages contain only G. duodenalis from animals (assemblages C and E, and undefined cat and rat genotypes). One of the assemblages in humans, assemblage B, also contains all beaver isolates and some isolates from muskrats, rabbits, and mice, which strongly suggests that these animal isolates have the potential to infect humans. Giardia from beavers has been suggested as the source of infection for backpackers and some waterborne outbreaks of giardiasis (27,30). Results of our study provide genetic evidence to substantiate these claims.
The TPI-based genotyping tool is also useful in epidemiologic investigations of giardiasis in humans (15,45,46). A recent study in the United Kingdom of sporadic cases of human giardiasis used a TPI-based PCR–restriction fragment length polymorphism genotyping tool. Of the 33 TPI-PCR–positive infected patients, 21 (64%) were infected with assemblage B, 9 (27%) with assemblage A, and 3 (9%) samples were mixed infections of assemblages A and B (47). Similar results were obtained with samples from a nursery outbreak, in which 21 (88%) of 24 samples were shown to be G. duodenlais assemblage B parasites; the rest were assemblage A parasites (47). The intragenotypic variations of TPI in assemblage B identified in the present study should be useful in subtyping outbreak isolates. Because Giardia spp. have a clonal population structure (40), the use of a typing system based on sequence analysis of a single genetic locus with high sequence heterogeneity, such as TPI, can provide a resolution as high as multilocus sequence typing.
The results of our study suggest that the TPI gene is a good phylogenetic marker for analysis of the molecular evolutionary and taxonomic relationship of G. duodenalis parasites. The genetic relationship shown by phylogenetic analysis of the TPI gene is largely in agreement with that obtained at other genetic loci. Results of the molecular analyses support the conclusion that G. duodenalis is a species-complex, a finding that should be useful in the revision of Giardia taxonomy and standardization of Giardia nomenclatures. Results of this study also indicate that Giardia parasites from beavers, muskrats, mice, and rabbits represent a potential public health concern.
Dr. Sulaiman is a guest researcher in the Division of Parasitic Diseases, National Center for Infectious Diseases, Centers for Disease Control and Prevention. His major interests focus on the molecular epidemiology and phylogenetics of protozoan parasites.
Acknowledgment
We thank Padma Vijyalakshmi and William Wong for providing specimens and Kristie Ludwig and Robert Palmer for technical support.
References
- Thompson RCA, Hopkins RA, Homan WL. Nomenclature and genetic groupings of Giardia infecting mammals. Parasitol Today. 2000;16:210–8. DOIPubMedGoogle Scholar
- Filice FP. Studies on the cytology and life history of a Giardia from the laboratory rat. Univ Calif Publ Zool. 1952;57:53–46.
- Erlandsen SL, Bemrick WL. SEM evidences for a new species, Giardia psittaci. J Parasitol. 1987;73:623–9. DOIPubMedGoogle Scholar
- Erlandsen SL, Bemrick WJ, Wellis CL, Feely DE, Kundson L, Cambell SR, Axenic culture and characterization of Giardia ardeae from the great blue heron (Ardea herodias). J Parasitol. 1990;76:717–24. DOIPubMedGoogle Scholar
- Feely DE. Morphology of the cyst of Giardia microti by light and electron microscopy. J Protozool. 1988;35:52–4.PubMedGoogle Scholar
- van Keulen H, Feely DE, Macechko T, Jarrol EL, Erlandsen SL. The sequence of Giardia small subunit rRNA shows that voles and muskrats are parasitized by a unique species, Giardia microti. J Parasitol. 1998;84:294–300. DOIPubMedGoogle Scholar
- Campbell SR, van Keulen H, Erlandsen SL, Senturia JB, Jarrol EL. Giardia sp: Comparison of electrophoretic karyotypes. Exp Parasitol. 1990;71:470–82. DOIPubMedGoogle Scholar
- van Keulen H, Gutell R, Gates M, Campbell S, Erlandsen SL, Jarrol EL, Unique phylogenetic position of Diplomonadida based on the complete small subunit ribosomal RNA sequence of Giardia ardeae, Giardia muris, Giardia duodenalis and Hexamita sp. FASEB J. 1993;7:223–31.PubMedGoogle Scholar
- Homan WL, van Enckevort FHJ, Limper L, van Eys GJJM, Schoone GJ, Kasprzak W, Comparison of Giardia isolates from different laboratories by isoenzyme analysis and recombinant DNA probes. Parasitol Res. 1992;78:316–23. DOIPubMedGoogle Scholar
- Nash TE, Mowatt MR. Identification and characterization of a Giardia lamblia group-specific gene. Exp Parasitol. 1992;75:369–78. DOIPubMedGoogle Scholar
- Maryhofer G, Andrews RH, Ey PL, Chilton NB. Division of Giardia isolates from humans into two genetically distinct assemblages by electrophoretic analysis of enzymes coded at 27 loci and comparison with Giardia muris. Parasitology. 1995;111:11–7. DOIPubMedGoogle Scholar
- Monis PT, Andrews RH, Mayrhofer G, Ey PL. Molecular systematics of the parasitic protozoan Giardia intestinalis. Mol Biol Evol. 1999;16:1135–44.PubMedGoogle Scholar
- Nash TE, McCutchan T, Keister D, Dame JB, Conard JD, Gillin FD. Restriction endonuclease analysis of DNA from 15 Giardia isolates obtained from humans and animals. J Infect Dis. 1985;152:64–73.PubMedGoogle Scholar
- Thompson RCA. Giardiasis as a re-emerging infectious disease and its zoonotic potential. Int J Parasitol. 2000;30:1259–67. DOIPubMedGoogle Scholar
- Baruch AC, Isaac-Renton J, Adam RD. The molecular epidemiology of Giardia lamblia: a sequence-based approach. J Infect Dis. 1996;174:233–6.PubMedGoogle Scholar
- Ey PL, Andrews RH, Maryhofer G. Differentiation of major genotypes of Giardia intestinalis by polymerase chain reaction analysis of a gene encoding a trophozoite surface antigen. Parasitology. 1993;106:347–56. DOIPubMedGoogle Scholar
- Monis PT, Mayrhofer G, Andrews RH, Homan WL, Limper L, Ey PL. Molecular genetic analysis of Giardia intestinalis isolates at the glutamate dehydrogenase locus. Parasitology. 1996;112:1–12. DOIPubMedGoogle Scholar
- Genetics Computer Group. Wisconsin Package Version 90. Madison (WI): Genetics Computer Group; 1996.
- Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–25.PubMedGoogle Scholar
- van de Peer Y, De Wachter R. TREECON for Windows: a software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Comput Appl Biosci. 1994;10:569–70.PubMedGoogle Scholar
- Felsenstein J. Confidence limits on the phylogenies: an approach using bootstrap. Evolution. 1985;39:783–91. DOIGoogle Scholar
- Efron B, Halloran E, Holmes S. Bootstrap confidence levels for phylogentic trees. Proc Natl Acad Sci U S A. 1996;93:13429–34. DOIPubMedGoogle Scholar
- Barta JR. Investigating phylogenetic relationships within the Apicomplexa using sequence data: the search of homology. Methods. 1997;13:81–8. DOIPubMedGoogle Scholar
- Monis PT. The importance of systematics in parasitological research. Int J Parasitol. 1999;29:381–8. DOIPubMedGoogle Scholar
- Olson ME, McAllister TA, Deselliers L, Morck DW, Cheng KJ, Buret AG, Effects of giardiasis on production in a domestic ruminant (lamb) model. Am J Vet Res. 1995;56:1470–4.PubMedGoogle Scholar
- Wallis PM, Buchanan-Mappin JM, Faubert GM, Belosevic M. Reservoirs of Giardia spp. in southwestern Alberta. J Wildl Dis. 1984;20:279–83.PubMedGoogle Scholar
- Pacha RE, Clark GW, Williams EA. Occurrence of Campylobacter jejuni and Giardia species in muskrats (Ondarta zibethica). Appl Environ Microbiol. 1985;50:177–8.PubMedGoogle Scholar
- Kirkpatrick CE, Benson CE. Presence of Giardia spp. and absence of Salmonella spp. in New Jersey muskrats (Ondarta zibethicus). Appl Environ Microbiol. 1987;53:1790–2.PubMedGoogle Scholar
- Monzingo DL, Hibler CP. Prevalence of Giardia sp. in a beaver colony and the resulting environmental contamination. J Wildl Dis. 1987;23:576–85.PubMedGoogle Scholar
- Patton S, Rabinowitz AR. Parasites of wild felidae in Thiland: a coprological survey. J Wildl Dis. 1994;30:472–5.PubMedGoogle Scholar
- Karanis P, Opiela K, Renoth S, Seith HM. Possible contamination of surface waters with Giardia spp. through muskrats. Zentralbl Bakteriol. 1996;284:302–6.PubMedGoogle Scholar
- Nizeyi JB, Mwebe R, Nanteza A, Cranfield MR, Kalema GR, Graczyk TK. Cryptosporidium sp. and Giardia sp. infectious in mountain gorillas (Gorilla gorilla beringi) of the Bwindi Impenetrable National Park, Uganda. J Parasitol. 1999;85:1084–8. DOIPubMedGoogle Scholar
- Rickard LG, Siefker C, Boyle CR, Gentz EJ. The prevalence of Cryptosporidium and Giardia spp. in fecal samples from free-ranging white-tailed deer (Odocoileus virginianus) in the southeastern United States. J Vet Diagn Invest. 1999;11:65–72.PubMedGoogle Scholar
- Visvesvara GS, Dickerson JW, Healy GR. Variable infectivity of human-derived Giardia lamblia cysts for Mongolian gerbils (Meriones unguiculatus). J Clin Microbiol. 1988;26:837–41.PubMedGoogle Scholar
- Hall ML, Costa ND, Thompson RCA, Lymbery AJ, Meloni BP, Wales RG. Genetic variants of Giardia duodenalis differ in their metabolism. Parasitol Res. 1992;78:712–4. DOIPubMedGoogle Scholar
- Andrews RH, Chilton NB, Maryhofer G. Selection of specific genotypes of Giardia intestinalis by growth in vitro and in vivo. Parasitology. 1992;105:375–86. DOIPubMedGoogle Scholar
- Binz N, Thompson RCA, Lymbery AJ, Hobbs RP. Comparative studies on the growth dynamics of two genetically distinct isolates of Giardia duodenalis in vitro. Int J Parasitol. 1992;22:195–202. DOIPubMedGoogle Scholar
- Andrews RH, Adams M, Boreham PFL, Maryhofer G, Meloni BP. Giardia intestinalis: electrophoretic evidence for a species-complex. Int J Parasitol. 1989;19:183–90. DOIPubMedGoogle Scholar
- Meloni BP, Lymbery AJ, Thompson RCA. Genetic characterization of isolates of Giardia duodenalis by enzyme electrophoresis: Implication of reproductive biology population structure taxonomy and epidemiology. J Parasitol. 1995;81:368–83. DOIPubMedGoogle Scholar
- Ey PL, Mansouri M, Kulda J, Nohynkova E, Monis PT. Genetic analysis of Giardia from hoofed farm animals reveals artiodactyls-specific and potentially zoonotic genotypes. J Eukaryot Microbiol. 1997;44:626–35. DOIPubMedGoogle Scholar
- Monis PT, Andrews RH, Mayrhofer G, Kulda J, Isaac-Renton JL, Ey PL. Novel lineages of Giardia intestinalis identified by genetic analysis of organisms isolated from dogs in Australia. Parasitology. 1998;116:7–19. DOIPubMedGoogle Scholar
- van Keulen H, Campbell SR, Erlandsen SL, Jarrol L. Cloning and restriction enzyme mapping of ribosomal DNA of Giardia duodenalis, Giardia ardeae and Giardia muris. Mol Biochem Parasitol. 1991;46:275–84. DOIPubMedGoogle Scholar
- Mowatt MR, Weinbach EC, Howard TC, Nash TT. Complementation of Escherichia coli glycolysis mutant by Giardia lamblia triosephosphate isomerase. Exp Parasitol. 1994;78:85–92. DOIPubMedGoogle Scholar
- Lu S, Li J, Zhang Y, Wen J, Wang F. The intraspecific difference of the triose phosphate isomerase (tim) gene from Giardia lamblia. Chin Med J (Engl). 2002;115:763–6.PubMedGoogle Scholar
- Lu S, Wen J, Li J, Wang F. DNA sequence analysis of the triose phosphate isomerase gene from isolates of Giardia lamblia. Chin Med J (Engl). 2002;115:99–102.PubMedGoogle Scholar
- Amar CFL, Dear PH, Pedraza-Diaz S, Looker N, Linnane E, McLauchlin J. Sensitive PCR-restriction fragment length polymorphism assay for detection and genotyping of Giardia duodenalis in human feces. J Clin Microbiol. 2002;40:446–52. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 9, Number 11—November 2003
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Lihua Xiao, Division of Parasitic Diseases, National Center for Infectious Diseases, Centers for Disease Control and Prevention, Building 22, Mailstop F12, 4770 Buford Highway, Atlanta, GA 30341-3717, USA; fax: (770) 488-4454
Top