Volume 14, Number 7—July 2008
Research
Transmission of Bartonella henselae by Ixodes ricinus
Cite This Article
Citation for Media
Abstract
Bartonella spp. are facultative intracellular bacteria associated with several emerging diseases in humans and animals. B. henselae causes cat-scratch disease and is increasingly associated with several other syndromes, particularly ocular infections and endocarditis. Cats are the main reservoir for B. henselae and the bacteria are transmitted to cats by cat fleas. However, new potential vectors are suspected of transmitting B. henselae, in particular, Ixodes ricinus, the most abundant ixodid tick that bites humans in western Europe. We used a membrane-feeding technique to infect I. ricinus with B. henselae and demonstrate transmission of B. henselae within I. ricinus across developmental stages, migration or multiplication of B. henselae in salivary glands after a second meal, and transmission of viable and infective B. henselae from ticks to blood. These results provide evidence that I. ricinus is a competent vector for B. henselae.
Bartonella spp. are facultative intracellular bacteria associated with several emerging diseases in humans and animals (1). Domestic animals and wildlife represent a large reservoir for Bartonella spp., and at least 10 species or subspecies have been reported to cause zoonotic infections. B. henselae causes cat-scratch disease, possibly the most common zoonosis acquired from domestic animals in industrialized countries and is becoming increasingly associated with other syndromes, particularly ocular infections and endocarditis (2–6). Although cat fleas are well-established vectors for B. henselae (7–10), transmission by other arthropods, in particular ticks, has been suggested (11–13). Ixodes ricinus is the most widespread and abundant ixodid tick in western Europe and is frequently associated with bites in humans. It is a vector of emerging zoonotic pathogens including Borrelia burgdorferi sensu lato (14), Anaplasma phagocytophilum (15), and Babesia spp (16).
Direct proof of transmission of Bartonella spp. by a tick was reported by Noguchi in 1926 (17), who described experimental transmission of B. bacilliformis (cause of Oroya fever) to monkeys by Dermacentor andersoni. In this study, ticks were allowed to feed on infected monkeys for 5 days. After removal, partially engorged ticks were placed on healthy monkeys in which disease then developed. This study showed that ticks could acquire and transmit the bacteria but did not demonstrate their vector competence or transtadial transmission throughout the tick’s life cycle. Since this early study, the role of ticks in Bartonella spp. transmission has been strongly implied but never definitively demonstrated. Bartonella spp. DNA was detected in questing and engorged nymphs and adults Ixodes spp. collected in North America, Europe, and Asia (13,18–26). If one considers that ixodid ticks feed only once per stage, Bartonella spp. DNA in questing ticks suggests transtadial transmission of these bacteria.
Other observations support Bartonella spp. transmission by ticks. Co-occurrence of Bartonella spp. with known tick-borne pathogens such as B. burgdorferi sensu lato, A. phagocytophilum, or Babesia spp. is not a rare event in ticks and hosts (13,19,24,27). A study conducted in a veterinary hospital in the United States (California) demonstrated that all dogs with endocarditis and infected with Bartonella spp. were also seropositive for A. phagocytophilum (28). In humans, several case studies have reported patients with concurrent Bartonella seropositivity and detection of Bartonella spp. DNA in their blood, along with B. burgdorferi infection of the central nervous system after tick bites (11,29). Moreover, Bartonella spp. DNA has been detected in human blood cells after a tick bite (30), and 3 patients with B. henselae bacteremia who had no history of contact with cats but had sustained tick bites were reported in Texas (12). Finally, tick exposure was determined to be a risk factor associated with B. vinsonii seropositivity in dogs (31).
Because Bartonella spp. are emerging human pathogens and Ixodes spp. can transmit a large spectrum of pathogens to humans, the capability of Ixodes spp. in transmitting human pathogenic Bartonella spp. should be determined. We used a membrane-feeding technique to infect I. ricinus with B. henselae, and investigated transtadial and transovarial transmission of viable and infective bacteria and putative transmission from tick saliva to blood during artificial blood meals.
Ticks
I. ricinus ticks were collected from the forest of Gâvre (Loire-Atlantique, France) in 2006 by flagging vegetation as described (32). Ticks were reared and maintained in chambers with a relative humidity of 80%–90% at 22°C before feeding on artificial skin. A total of 217 whole ticks were tested for Bartonella DNA.
Culturing of B. henselae
B. henselae (Houston-1 ATCC 49 882) were grown on 5% defibrinated sheep blood Columbia agar (CBA) plates, which were incubated at 35°C in an atmosphere of 5% CO2. After 10 days, bacteria were harvested, suspended in sterile phosphate-buffered saline (PBS), and used immediately for artificial feeding of ticks.
Housing of Cats
Three healthy male European cats were used (12–14 months of age, weight = 3.2–4.6 kg at the beginning of the study). These cats, which were bred by Harlan (Indianapolis, IN, USA), were imported at 9 months of age. Before the study, animals underwent clinical examinations and showed no signs of disease. Absence of Bartonella spp. was confirmed by blood culture, tests for DNA of Bartonella spp., and serologic analysis. Animals were housed singly in cages in compliance with European guidelines (cage surface 6,000 cm2, cage height 50 cm). Animals were allowed to acclimate for 3 months to the facility, diet, and handling before the first blood sample was taken and subsequent infection. Animals were fed ad libitum with standard feline maintenance diet (Harlan) and received water with no restrictions. Animal care was provided in accordance with the good animal care practice.
Feeding of I. ricinus Ticks with Ovine Blood
Figure 1
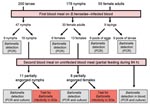
Figure 1. Experimental framework of Ixodes ricinus tick infection by Bartonella henselae–infected blood. Ticks (200 larvae, 178 nymphs, and 55 female adults) were engorged by feeding through artificial skin on B. henselae–infected blood...
The general experimental framework of artificial feeding is shown in Figure 1. Ovine blood used in all experiments was obtained from 3 sheep reared at the National Veterinary School in Nantes, France. Absence of Bartonella spp. in the blood of these sheep was confirmed by culture assay and tests for DNA of Bartonella spp. by PCR. Lithium heparin–coated vacutainer tubes (Becton Dickinson, Le Pont de Claix-Cedex, France) were used to draw blood by venipuncture. Blood was depleted of functional complement by heat treatment (incubation for 30 min at 56°C) before use. To avoid fungal and bacterial contamination during feeding experiments, decomplemented blood was supplemented with fosfomycin (20 μg/mL) and amphotericin B (0.25 μg/mL), which were previously determined to have no effect on B. henselae viability.
Feeding of Ticks with B. henselae–Infected Blood
The method of artificial feeding used in this study was adapted from the method of Bonnet et al. (33). Ticks were placed in 75-cm2 tissue culture flasks pierced at the top to accommodate a 4-cm diameter glass feeder. The feeder apparatus was closed with a parafilm membrane at the top and a gerbil or rabbit skin membrane at the bottom. Gerbil skins were used for feeding larvae and rabbit skins for feeding nymphs and adults (33). To attract ticks, a constant temperature (37°C) was maintained by a water-jacket circulation system through the glass feeder. For blood infection, 10 μL of B. henselae suspension at a concentration of 109 CFU/mL in PBS was added to 10 mL of blood supplemented with fosfomycin and amphotericin B. The culture box containing ticks was placed under the feeding apparatus, and 3 mL of B. henselae–infected ovine blood, changed twice a day, was introduced until the ticks were replete. To feed adult ticks, an equal number of males (for reproduction) and females were used. Separate apparatuses were used to engorge 200 larvae, 178 nymphs, and 55 female adults. As a control, 50 nymphs were fed under the same conditions on uninfected decomplemented ovine blood.
After feeding, larvae and nymphs were allowed to molt to nymphs and adults, respectively, and engorged females were allowed to lay eggs. Between each feeding on skin, ticks were starved for at least 2 months.
Feeding with Uninfected Blood of Ticks Infected with B. henselae at Preceding Stages
Nymphs and adults fed on blood infected with B. henselae at preceding stages were fed with uninfected, decomplemented, ovine blood in 2 glass feeders as described above. After 84 h of refeeding, nymphs and females attached to the skin were removed and dissected. Bartonella spp. DNA was detected every 24 h in blood from the first 48 h of attachment onward. At each time point, 3 mL of blood was removed and centrifuged for 30 min at 3,000 × g. The supernatant was aspirated, and the pellet (200 μL) was used to detect bacterial DNA. After 84 h of feeding, 10 μL of blood was used for B. henselae culture.
Tick Dissection
Salivary glands (SGs) from I. ricinus adults and nymphs were dissected under a magnifying glass in sterile PBS. All dissection material was cleaned with DNA-off (Eurobio, Courtaboeuf, France) and rinsed with sterile water between each sample. Individual pairs of tick SGs and the remaining tick carcasses were suspended in 150 μL of PBS before culture and used to infect cats or frozen at –80°C until DNA extraction.
Detection of B. henselae in Tick and Blood Samples
DNA Extraction
Carcasses (ticks without SGs), entire ticks, and pools of 50 larvae were mechanically disrupted as described (34). Eggs were crushed by using a microtissue grinder and dissected SGs were directly used for DNA extraction. DNA was extracted from all tick samples by using the Nucleospin Tissue kit according to the manufacturer’s instructions (Macherey-Nagel, Duren, Germany). DNA was extracted from blood samples by using the Nucleospin Blood Quick Pure kit (Macherey-Nagel). For carcasses and entire ticks, the final elution volume was 100 μL for adults and 30 μL for nymphs. For DNA from SGs, the final elution volume was 50 μL for adults and 20 μL for nymphs. Pools of eggs, larvae, and blood DNA were eluted in volumes of 50 μL.
PCR Amplification
Efficiency of tick DNA extraction was evaluated in all samples by amplification of a fragment of the tick mitochondrial 16S rRNA gene by using tick-specific primers TQ16S+1F (5′-CTGCTCAATGATTTTTTAAATTGCTGTGG-3′) and TQ16S-2R (5′-ACGCTGTTATCCCTAGAG-3′) as described (35). A seminested PCR was used to detect B. henselae DNA. Amplification was initially performed with 5 μL of DNA extract and universal bacteria primers amplifying a 535-bp fragment of the 16S rRNA gene: pc535 (5′-GTATTACCGCGGCTGCTGGCA-3′) and p8 (5′AGAGTTTGATCCTGGCTCAG-3′).
The second seminested amplification was performed with 5 μL of a 100-fold dilution of the first PCR product and primers pc535 and bsp16F (5′-TCTCTACGGAATAACACAGA-3′) a 16S rRNA Bartonella spp.–specific primer, and resulted in amplification of a 337-bp product. The PCR cycle was identical for both amplification reactions: an initial denaturation step for 8 min at 94°C; 35 cycles of denaturation for 1 min at 94°C, annealing for 1 min at 54°C, and extension for 1 min at 72°C; and a final extension step at 72°C for 10 min. Each reaction was conducted in a total volume of 25 μL with 0.5 μmol/μL of each primer, 2.5 mmol/L of each dNTP, 2.5 μL of 10× PCR buffer, and 1 U of Taq DNA polymerase (Takara Biomedical Group, Shiga, Japan). Negative (ticks fed on uninfected ovine blood) and positive (B. bacilliformis DNA to easily detect any cross-contamination) controls were included in each assay. All PCRs were performed in a thermocycler MyCycler (Bio-Rad, Strasbourg, France).
Sequencing and Sequence Analysis
The expected 337-bp PCR product was isolated by agarose gel electrophoresis, excised from the gel, purified by using NucleoSpin Extract II (Macherey-Nagel), and sent to QIAGEN (Hilden, Germany) for direct sequencing. Sequences were compared with known sequences listed in the GenBank nucleotide sequence databases by using the BLAST search option of the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov/BLAST).
B. henselae Culture from Tick SGs and Blood
After refeeding and dissection, 4 pairs of SGs from 4 potentially infected adults females and 3 nymphs were incubated in 1 mL of Schneider Drosophila medium (Invitrogen, Cergy-Pontoise, France) at 35°C in an atmosphere of 5% CO2. After 6 days of incubation, 10-μL samples were placed on CBA plates. Ten microliters of blood was removed from the glass feeder after 84 h of refeeding with adults suspected of having B. henselae and incubated for 6 days in Schneider Drosophila medium before being placed on CBA plates.
Infection of Cats with Adult and Nymph SGs Potentially Infected with B. henselae
One cat was intravenously infected with 150 μL of PBS containing SGs from refed adult ticks suspected of having B. henselae; another cat was infected with 150 μL of PBS. Before handling, cats were anesthetized with ketamine (10–15 mg/kg bodyweight) and diazepam (100 μg/kg bodyweight) given intravenously. One milliliter of blood was obtained from the jugular vein at days 7, 14, and 21 postinfection, and 100 μL of blood dilutions (1:10, 1:100, and 1:1,000) was directly placed on sheep blood CBA plates. Colonies were counted and CFU/mL was estimated after incubation for 10 days at 35°C in an atmosphere of 5% CO2.
Detection of Bartonella spp. DNA in Questing I. ricinus Ticks
We tested 217 ticks collected in the forest of Gavre to determine prevalence of Bartonella spp. DNA. None of the 98 nymphs, 49 female ticks, and 70 male ticks collected in the forest showed amplification of the Bartonella spp. 16S rRNA gene.
Transstadial Transmission of B. henselae DNA by I. ricinus Ticks
A total of 433 I. ricinus ticks were used in 2 independent experiments with feeding on artificial skin. Of these ticks, 169 (84.5%) of 200 larvae, 111 (62.3%) of 178 nymphs, and 19 (34.5%) of 55 female adults were successfully engorged and spontaneously detached. After feeding and detachment, all ticks were maintained in humidity chambers to enable molting or laying of eggs. After 3 months, 47 larvae (27.8%) and 58 nymphs (30.1%) molted into nymphs and adults (28 males and 30 females), respectively. Of the 19 engorged females, 9 laid eggs.
Figure 2
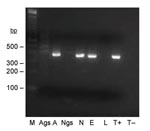
Figure 2. Seminested PCR detection of Bartonella spp. DNA in Ixodes ricinus ticks fed on B. henselae–infected ovine blood at preceding stage. Lane M, 100-bp DNA molecular mass marker; lane Ags, salivary glands...
Nine nymphs exposed to infection as larvae were tested for B. henselae DNA. All carcasses showed amplification of the expected 337-bp DNA fragment, and no amplification product was obtained from SGs. Four carcasses of 6 females exposed to B. henselae as nymphs showed the 337-bp DNA fragment, and no amplification product was obtained with SGs (Table, Figure 2). No amplified fragment was detected with DNA extracts from SGs or carcasses of control ticks fed on uninfected blood. All PCR products were sequenced and shared 100% identity with the 16S rRNA gene of B. henselae Houston-1 (Genbank accession no. BX897699).
Transovarial Transmission of B. henselae DNA by I. ricinus Ticks
Among 9 pools of eggs laid by females fed on B. henselae–infected blood, 3 showed amplification of the expected Bartonella spp.–specific 337-bp DNA fragment. No amplification was obtained with larvae from eggs positive or negative for Bartonella spp. DNA (Figure 2).
Transmission of Viable and Infective B. henselae by I. ricinus Ticks
Figure 3
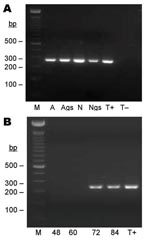
Figure 3. Seminested PCR detection of Bartonella spp. DNA after partial refeeding of infected ticks. A) Bartonella spp. DNA detection in Ixodes ricinus ticks fed on B. henselae–infected blood at previous development stages...
Eighteen nymphs and 13 female adults fed on B. henselae–infected blood at preceding life stages were refed for 84 h with uninfected blood. DNA extracts were prepared from SGs and carcasses of 7 partially engorged nymphs and 3 partially engorged females. Bartonella spp. DNA was amplified from carcasses and SGs of 4 nymphs and 1 female. For 1 nymph and 1 female, the specific 337-bp DNA fragment was amplified only in SG DNA extracts but not in carcass extracts (Table; Figure 3, panel A). Four pairs of SGs from 4 partially engorged females and 3 pairs of SGs from 3 partially engorged nymphs were incubated separately in Schneider Drosphila medium for 6 days before being placed on sheep blood agar. B. henselae colonies appeared after 7 days for all SGs tested, which indicated the presence of viable bacteria in SGs from adults and nymphs.
To determine whether B. henselae in SGs of nymphs and adults were infective, we infected 2 healthy cats with 1 pair of SGs from a nymph and 1 pair of SGs from a female adult. The cat infected with SGs from an adult became bacteremic 7 days after infection (5 × 105 CFU/mL at day7, 6 × 106 CFU/mL at day 14, and 1 × 106 CFU/mL at day 21), and the cat infected with SGs from a nymph became bacteremic after 14 days of infection (8 × 105 CFU/mL at day 14 and 2 × 107 CFU/mL at day 21).
Blood samples from feeders were obtained every 12 h for the first 48 h to detect B. henselae. In the first 60 h, B. henselae DNA or viable B. henselae was not detected in blood samples. After 72 h of refeeding of infected ticks, Bartonella spp. DNA was successfully amplified (Figure 3, panel B), and B. henselae colonies were isolated from these blood samples after preincubation for 6 days in Schneider Drosophila medium, which indicated viability of the transmitted bacteria.
This study demonstrated transmission of B. henselae by I. ricinus ticks across different developmental stages, migration and multiplication of viable and infective B. henselae in SGs after a second blood meal, and transmission of B. henselae from ticks to blood. These findings indicate that I. ricinus is a competent vector for B. henselae.
Vector biologists and epidemiologists have suggested that ticks may play a role in transmission of Bartonella spp (11,12,19,23,25,28,29). This suggestion was based on indirect data for detection of bacterial DNA in ticks (18,19,24), humans exposed to tick bites (30), or serologic evidence of co-infection of humans with pathogens known to be transmitted by ticks (11,36). Difficulties in rearing I. ricinus and lack of a rodent model for B. henselae infection may explain the absence of data demonstrating the role of this tick as a vector of B. henselae. Recent development of an artificial method suitable for feeding ticks (33) enabled us to study experimental infection of ticks with blood containing B. henselae, to monitor B. henselae through various tick stages, and to evaluate putative transmission of bacteria from the tick to blood.
To select a tick population with the lowest Bartonella spp. DNA prevalence, we estimated the prevalence of Bartonella spp. DNA in questing I. ricinus collected in different areas in France. The lowest Bartonella spp. DNA prevalence was in Loire-Atlantique (19 and unpub. data). We thus used ticks collected in this area for our study.
We detected B. henselae DNA in 100% of carcasses from nymphs and 67% of carcasses from adults fed on ovine blood containing B. henselae at their preceding stages. No B. henselae DNA was amplified in corresponding SGs in a nested PCR, which is more sensitive than amplification of the classic citrate synthase gene. This result demonstrated that bacteria could be ingested by I. ricinus larvae and nymphs during feeding on artificial skin and that bacterial DNA was maintained in the tick after molting. However, no or undetectable numbers reached the SGs.
Although bacterial DNA was detected in eggs laid by females fed on blood containing B. henselae, larvae obtained from these eggs were PCR-negative for B. henselae. This finding suggests external contamination of eggs with DNA rather than transovarial persistence of bacteria.
When molted nymphs and female ticks potentially contaminated with B. henselae at their previous developmental stage were refed on uninfected blood, viable B. henselae were detected in SGs after 84 h of engorgement. Two hypotheses could explain the absence of detectable bacterial DNA in SGs after an infected blood meal and molting, when it becomes detectable after a partial refeeding blood meal. The first hypothesis is that the 84-h refeeding period may act as a stimulus and enable migration of bacteria from the gut to SGs of the tick, as previously described for B. burgdorferi sensu lato (14). The second hypothesis is that this refeeding period may stimulate multiplication of bacteria already present in SGs, but at undetectable levels. More investigations are needed to validate one of these hypotheses. Bacteria located in SGs of nymphs and adults are infective because injection of 1 pair of infected SGs into cats induced high levels of bacteremia. Cats became bacteremic in the first 2 weeks after injection, as described for cat infection with B. henselae by fleas, and bacteremia levels were similar to those observed in cats infected by flea bites (7,8).
Viable B. henselae in blood after 72 h of feeding of ticks with B. henselae–infected ticks demonstrated its transmission from the tick to the blood by mouthparts of the ticks. The duration of multiplication or migration described above would explain such a delay in bacterial transmission.
Because our results have demonstrated competence of I. ricinus for transmission of B. henselae, cat models of B. henselae transmission by ticks are needed to confirm that cats can be infected with B. henselae by tick bites. Further investigations are also needed to evaluate the capacity of I. ricinus to transmit B. henselae to cats and humans. Such transmission could occur because cats, although not common hosts for I. ricinus, can be infested with this tick. In France, attached I. ricinus are commonly found on cats brought to veterinarians (J. Guillot, pers. comm.). In Great Britain, Ogden et al. (37) reported cats with woodland and moorland habitats as hosts for I. ricinus. Podsiadly et al. (38) reported B. henselae in cats and in I. ricinus removed from those cats in Poland.
In conclusion, we demonstrated by using feeding on artificial skin that B. henselae, the cause of cat-scratch disease in humans, could be transmitted by ticks through saliva. Although further investigations are needed to clarify the epidemiology of such transmission, health authorities must take into account the possibility of bartonellosis in persons exposed to tick bites, and B. henselae must be identified as a tick-borne pathogen.
Ms Cotté is a doctoral student in microbiology at the Unité Mixte de Recherche 956 in Maisons-Alfort, France. Her research interest is transmission of Bartonella spp. by ticks.
Acknowledgments
We thank A. Agoulon and J.P. Delatre for collecting ticks.
This study was supported by Merial Laboratory, Lyon, France, and the French National Institute for Agronomic Science, Paris, France.
References
- Boulouis HJ, Chang CC, Henn JB, Kasten RW, Chomel BB. Factors associated with the rapid emergence of zoonotic Bartonella infections. Vet Res. 2005;36:383–410. DOIPubMedGoogle Scholar
- Anderson BE, Neuman MA. Bartonella spp. as emerging human pathogens. Clin Microbiol Rev. 1997;10:203–19.PubMedGoogle Scholar
- Oteo JA, Castilla A, Arosey A, Blanco JR, Ibarra V, Morano LE. Endocarditis due to Bartonella spp. Three new clinical cases and Spanish literature review [in Spanish]. Enferm Infecc Microbiol Clin. 2006;24:297–301. DOIPubMedGoogle Scholar
- Marsilia GM, La Mura A, Galdiero R, Galdiero E, Aloj G, Ragozzino A. Isolated hepatic involvement of cat scratch disease in immunocompetent adults: enhanced magnetic resonance imaging, pathological findings, and molecular analysis–two cases. Int J Surg Pathol. 2006;14:349–54. DOIPubMedGoogle Scholar
- Curi AL, Machado DO, Heringer G, Campos WR, Orefice F. Ocular manifestation of cat-scratch disease in HIV-positive patients. Am J Ophthalmol. 2006;141:400–1. DOIPubMedGoogle Scholar
- Chomel BB, Boulouis HJ, Maruyama S, Breitschwerdt EB. Bartonella spp. in pets and effect on human health. Emerg Infect Dis. 2006;12:389–94.PubMedGoogle Scholar
- Chomel BB, Kasten RW, Floyd-Hawkins K, Chi B, Yamamoto K, Roberts-Wilson J, Experimental transmission of Bartonella henselae by the cat flea. J Clin Microbiol. 1996;34:1952–6.PubMedGoogle Scholar
- Higgins JA, Radulovic S, Jaworski DC, Azad AF. Acquisition of the cat scratch disease agent Bartonella henselae by cat fleas (Siphonaptera:Pulicidae). J Med Entomol. 1996;33:490–5.PubMedGoogle Scholar
- Foil L, Andress E, Freeland RL, Roy AF, Rutledge R, Triche PC, Experimental infection of domestic cats with Bartonella henselae by inoculation of Ctenocephalides felis (Siphonaptera:Pulicidae). J Med Entomol. 1998;35:625–8.PubMedGoogle Scholar
- Finkelstein JL, Brown TP, O’Reilly KL, Wedincamp J Jr, Foil LD. Studies on the growth of Bartonella henselae in the cat flea (Siphonaptera:Pulicidae). J Med Entomol. 2002;39:915–9.PubMedGoogle Scholar
- Eskow E, Rao RV, Mordechai E. Concurrent infection of the central nervous system by Borrelia burgdorferi and Bartonella henselae: evidence for a novel tick-borne disease complex. Arch Neurol. 2001;58:1357–63. DOIPubMedGoogle Scholar
- Lucey D, Dolan MJ, Moss CW, Garcia M, Hollis DG, Wegner S, Relapsing illness due to Rochalimaea henselae in immunocompetent hosts: implication for therapy and new epidemiological associations. Clin Infect Dis. 1992;14:683–8.PubMedGoogle Scholar
- Holden K, Boothby JT, Kasten RW, Chomel BB. Co-detection of Bartonella henselae, Borrelia burgdorferi, and Anaplasma phagocytophilum in Ixodes pacificus ticks from California, USA. Vector Borne Zoonotic Dis. 2006;6:99–102. DOIPubMedGoogle Scholar
- De Silva AM, Fikrig E. Growth and migration of Borrelia burgdorferi in Ixodes ricinus ticks during blood feeding. Am J Trop Med Hyg. 1995;53:397–404.PubMedGoogle Scholar
- Raoult D, Roux V. Rickettsioses as paradigms of new or emerging infectious diseases. Clin Microbiol Rev. 1997;10:694–719.PubMedGoogle Scholar
- Gorenflot A, Moubri K, Precigout E, Carcy B, Schetters TP. Human babesiosis. Ann Trop Med Parasitol. 1998;92:489–501. DOIPubMedGoogle Scholar
- Noguchi H. Etiology of Oroya fever: V. The experimental transmission of Bartonella bacilliformis by ticks (Dermacentor andersoni). J Exp Med. 1926;44:729–34. DOIGoogle Scholar
- Chang CC, Chomel BB, Kasten RW, Romano V, Tietze N. Molecular evidence of Bartonella spp. in questing adult Ixodes pacificus ticks in California. J Clin Microbiol. 2001;39:1221–6. DOIPubMedGoogle Scholar
- Halos L, Jamal T, Maillard R, Beugnet F, Le Menach A, Boulouis HJ, Evidence of Bartonella sp. in questing adult and nymphal Ixodes ricinus ticks from France and co-infection with Borrelia burgdorferi sensu lato and Babesia sp. Vet Res. 2005;36:79–87. DOIPubMedGoogle Scholar
- Morozova OV, Cabello FC, Dobrotvorsky AK. Semi-nested PCR detection of Bartonella henselae in Ixodes persulcatus ticks from Western Siberia, Russia. Vector Borne Zoonotic Dis. 2004;4:306–9. DOIPubMedGoogle Scholar
- Kim CM, Kim JY, Yi YH, Lee MJ, Cho MR, Shah DH, Detection of Bartonella species from ticks, mites and small mammals in Korea. J Vet Sci. 2005;6:327–34.PubMedGoogle Scholar
- Schouls LM, van de Pol I, Rijpekema SG, Schot CS. Detection and identification of Ehrlichia, Borrelia burgdorferi sensu lato, and Bartonella species in Dutch Ixodes ricinus ticks. J Clin Microbiol. 1999;37:2215–22.PubMedGoogle Scholar
- Sanogo YO, Zeaiter Z, Caruso G, Merola F, Shpynov S, Brouqui P, Bartonella henselae in Ixodes ricinus ticks (Acari: Ixodida) removed from humans, Belluno province, Italy. Emerg Infect Dis. 2003;9:329–32.PubMedGoogle Scholar
- Adelson ME, Rao RV, Tilton RC, Cabets K, Eskow E, Fein L, Prevalence of Borrelia burgdorferi, Bartonella spp., Babesia microti, and Anaplasma phagocytophila in Ixodes scapularis ticks collected in northern New Jersey. J Clin Microbiol. 2004;42:2799–801. DOIPubMedGoogle Scholar
- Podsiadly E, Chmielewski T, Sochon E, Tylewska-Wierzbanowska S. Bartonella henselae in Ixodes ricinus ticks removed from dogs. Vector Borne Zoonotic Dis. 2007;7:189–92. DOIPubMedGoogle Scholar
- Rar VA, Fomenko NV, Dobrotvorsky AK, Livanova NN, Rudakova SA, Fedorov EG, Tickborne pathogen detection, Western Siberia, Russia. Emerg Infect Dis. 2005;11:1708–15.PubMedGoogle Scholar
- Solano-Gallego L, Bradley J, Hegarty B, Sigmon B, Breitschwerdt E. Bartonella henselae IgG antibodies are prevalent in dogs from southeastern USA. Vet Res. 2004;35:585–95. DOIPubMedGoogle Scholar
- MacDonald KA, Chomel BB, Kittleson MD, Kasten RW, Thomas WP, Pesavento P. A prospective study of canine infective endocarditis in northern California (1999–2001): emergence of Bartonella as a prevalent etiologic agent. J Vet Intern Med. 2004;18:56–64. DOIPubMedGoogle Scholar
- Podsiadly E, Chmielewski T, Tylewska-Wierzbanowska S. Bartonella henselae and Borrelia burgdorferi infections of the central nervous system. Ann N Y Acad Sci. 2003;990:404–6.PubMedGoogle Scholar
- Morozova OV, Chernousova NI, Morozov IV. Detection of the Bartonella DNA by the method of nested PCR in patients after tick bites in Novosibirsk region [in Russian]. Mol Gen Mikrobiol Virusol. 2005;4:14–7.PubMedGoogle Scholar
- Pappalardo BL, Correa MT, York CC, Peat CY, Breitschwerdt EB. Epidemiologic evaluation of the risk factors associated with exposure and seroreactivity to Bartonella vinsonii in dogs. Am J Vet Res. 1997;58:467–71.PubMedGoogle Scholar
- Vassalo M, Paul RE, Pereiz-Eid C. Temporal distribution of the annual nymphal stock of Ixodes ricinus ticks. Exp Appl Acarol. 2000;24:941–9. DOIPubMedGoogle Scholar
- Bonnet S, Jouglin M, Malandrin L, Becker C, Agoulon A, L’Hostis M, Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology. 2006;134:197–207. DOIPubMedGoogle Scholar
- Halos L, Jamal T, Vial L, Maillard R, Suau A, Le Menach A, Determination of an efficient and reliable method for DNA extraction from ticks. Vet Res. 2004;35:709–13. DOIPubMedGoogle Scholar
- Black WC IV, Piesman J. Phylogeny of hard- and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc Natl Acad Sci U S A. 1994;91:10034–8. DOIPubMedGoogle Scholar
- Kordick SK, Breitschwerdt EB, Hegarty BC, Southwick KL, Colitz CM, Hancock SI. Coinfection with multiple tick-borne pathogens in a Walker Hound kennel in North Carolina. J Clin Microbiol. 1999;37:2631–8.PubMedGoogle Scholar
- Ogden NH, Cripps P, Davison CC, Owen G, Parry JM, Timms BJ, The ixodid tick species attaching to domestic dogs and cats in Great Britain and Ireland. Med Vet Entomol. 2000;14:332–8. DOIPubMedGoogle Scholar
- Podsiadly E, Chmielewski T, Marczak R, Sochon E, Tylewska-Wierzbanowska S. Bartonella henselae in the human environment in Poland. Scand J Infect Dis. 2007;39:956–62. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 14, Number 7—July 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Muriel Vayssier-Taussat, Unité Mixte de Recherche, Biologique et Immunologie Moléculaires des Infections Parasitaires et Fongiques, Agence Francaise de Securite Sanitaire des Aliments, 23 Rue du General de Gaulle, Maisons-Alfort 94700, France; email:
Top