Volume 16, Number 6—June 2010
Research
Oseltamivir-Resistant Influenza Viruses A (H1N1) during 2007–2009 Influenza Seasons, Japan
Cite This Article
Citation for Media
Abstract
To monitor oseltamivir-resistant influenza viruses A (H1N1) (ORVs) with H275Y in neuraminidase (NA) in Japan during 2 influenza seasons, we analyzed 3,216 clinical samples by NA sequencing and/or NA inhibition assay. The total frequency of ORVs was 2.6% (45/1,734) during the 2007–08 season and 99.7% (1,477/1,482) during the 2008–09 season, indicating a marked increase in ORVs in Japan during 1 influenza season. The NA gene of ORVs in the 2007–08 season fell into 2 distinct lineages by D354G substitution, whereas that of ORVs in the 2008–09 season fell into 1 lineage. NA inhibition assay and M2 sequencing showed that almost all the ORVs were sensitive to zanamivir and amantadine. The hemagglutination inhibition test showed that ORVs were antigenetically similar to the 2008–09 vaccine strain A/Brisbane/59/2007. Our data indicate that the current vaccine or zanamivir and amantadine are effective against recent ORVs, but continuous surveillance remains necessary.
Influenza A and B viruses are major pathogens that represent a threat to public health with subsequent economic losses worldwide (1). Vaccination is the primary method for prevention; antiviral drugs are used mainly for prophylaxis and therapy. Currently, 2 classes of drugs, matrix 2 (M2) blockers and neuraminidase inhibitors (NAIs) are available, but M2 blockers such as amantadine and rimantadine are not commonly used because of the rapid generation of resistance and lack of efficacy against influenza B virus (2–4). The NAIs zanamivir and oseltamivir are widely used because of effects against influenza A and B viruses and a low frequency of resistance. NAI virus surveillance studies by several groups have demonstrated that <1% of viruses tested show naturally occurring resistance to oseltamivir as of 2007 (5–10), indicating limited human-to-human transmission of these viruses.
At the beginning of the 2007–08 influenza season, however, detection of a substantially increased number of oseltamivir-resistant influenza viruses A (H1N1) (ORVs) was reported, mainly in countries in Europe where the prevalence varies, with the highest levels in Norway (67%) and France (47%) (11–14). These viruses showed a specific NA mutation with a histidine-to-tyrosine substitution at the aa 275 position (N1 numbering, H275Y), conferring high-level resistance to oseltamivir. Most of these ORVs were isolated from NAI-untreated patients and retained similar ability of human-to-human transmission to oseltamivir-sensitive influenza viruses A (H1N1) (OSVs) (10,15). In response to public health concerns about ORVs, the World Health Organization (WHO) directed Global Influenza Surveillance Network laboratories to intensify NAI surveillance and announced regularly updated summaries of ORV data collected from each laboratory on its website (16). This site reported that the global frequency increased from 16% (October 2007–March 2008) to 44% (April 2008–September 2008) to 95% (October 2008–January 2009), indicating that ORVs have spread rapidly around the world.
Figure 1

Figure 1. Weekly cases of influenza and isolation of influenza viruses in the 2007–08 and 2008–09 seasons in Japan (as of July 2, 2009). The National Epidemiologic Surveillance of Infectious Diseases (NESID) Network...
Japan has the highest annual level of oseltamivir usage per capita in the world, comprising >70% of world consumption (10). Such high use of oseltamivir has raised concerns about emergence of OSVs with increased resistance to this drug. Moreover, in Japan, 2 recent influenza seasons were dominated by influenza viruses A (H1N1) (Figure 1). If a high prevalence of ORVs is observed, primary selection of oseltamivir treatment for influenza patients should be reconsidered. Thus, monitoring ORVs is a serious public health issue.
To estimate the frequency of ORVs and characterize these viruses, we analyzed 1,734 clinical samples isolated from the 2007–08 season and 1,482 isolates from the 2008–09 season by NA sequencing and/or NAI inhibition assay. The total frequencies were 2.6% in the 2007–08 season and 99.7% in the 2008–09 season, indicating that ORVs increased dramatically in Japan.
Virus Testing
Influenza sentinel clinics send clinical specimens to local public health laboratories for virus isolation. Several culture tissues, including MDCK, Caco-2, and LLC-MK2, are used for virus isolation. Without successful viral isolation, clinical specimens are analyzed directly. Influenza viruses were collected from all 47 prefectures in Japan for this study; 1,734 samples of influenza A (H1N1) were isolated during the 2007–08 season (September 2007–August 2008) and 1,482 samples of influenza A (H1N1) were isolated in the 2008–09 season (September 2008–April 2009). During the 2007–08 season, viruses were isolated primarily after December 2007. All influenza viruses A (H1N1) were subjected to full or partial (nt 615–1076) NA sequencing to detect H275Y substitution on the N1 NA protein. Representative influenza viruses A (H1N1), including ORVs and OSVs, were subjected to NA inhibition assay (number of tested viruses isolated during the 2007–08 and 2008–09 seasons was 306 and 58, respectively), full NA sequencing (891 and 83), hemagglutinin (HA) 1 sequencing (299 and 83), M2 sequencing (288 and 79), and hemagglutinin inhibition (HI) test (187 and 59).
Sequence Analysis
The phylogenetic tree of NA and HA1 genes was constructed by neighbor-joining methods. The phylogenetic tree was described by representative ORVs and OSVs isolated from several prefectures in Japan. Sequence information for isolates from other countries was obtained from the Global Initiative on Sharing Avian Influenza Data and the Los Alamos National Laboratory database. All amino acid positions in the phylogenetic tree were described by N1 numbering.
NA Inhibition Assay
The chemiluminescent NA inhibition assay was performed by using the NA Star Kit (Applied Biosystems, Tokyo, Japan) with slight modifications of the instructions provided by the manufacturer. The final drug concentration ranged from 0.03 nmol/L to 6,500 nmol/L for oseltamivir and from 0.03 nmol/L to 12,500 nmol/L for zanamivir. Chemiluminescent light emission was measured by using an LB940 plate reader (Berthhold Technologies, Bad Wildbad, Germany). Drug concentrations required to inhibit NA activity by 50% (IC50) were calculated by a 4-parameter method using MikroWin 2000 version 4 software (Mikrotek Laborsysteme GmbH, Overath, Germany).
Hemagglutination Inhibition Test
The HI test was performed to evaluate the reactivity of ferret antiserum against 2008–09 vaccine strain A/Brisbane/59/2009, as described by the WHO manual (17). Antiserum was treated by receptor-destroying enzyme II (Denka Seiken, Tokyo, Japan) and adsorbed with packed turkey erythrocytes before testing to prevent nonspecific reaction. A 0.5% suspension of turkey erythrocytes was used for the HI test. Viruses with >8-fold reduced HI titer to the homologous titer of A/Brisbane/59/2009 antiserum were regarded as antigenic variants.
Statistical Analysis
To determine the cutoff value between NAI-resistant (outlier) and -sensitive viruses, box-and-whisker plots were used. The cutoff value was defined as upper quartile + 5.0×interquartile range from the 25th to 75th percentile. In this study, ORVs with H275Y were excluded from the overall population for statistical analysis. Outliers were excluded from the calculation of mean values and standard deviations for IC50.
Geographic Distribution of ORVs during the 2007–08 and 2008–09 Influenza Seasons
Figure 2
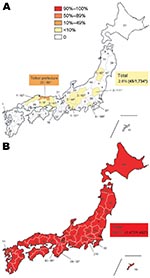
Figure 2. Geographic distribution of oseltamivir-resistant influenza viruses A (H1N1) (ORVs) with H275Y in Japan during the 2007–08 and 2008–09 seasons. The total number of influenza A (H1N1) isolates tested is described inside...
To estimate the frequency of influenza A (H1N1) ORVs in each prefecture of Japan, 1,734 isolates during the 2007–08 season and 1,482 isolates during the 2008–09 season were collected from all prefectures and examined by NA sequencing to detect the H275Y mutation in NA protein. In the 2007–08 season, 45 viruses possessing H275Y mutation (total frequency of ORVs 2.6%; Figure 2, panel A) were observed in 10 prefectures, indicating that the frequency of ORVs was significantly lower than that in countries in Europe and the United States (8,11–14). In Tottori prefecture, however, 22 of 68 influenza viruses A (H1N1) tested possessed H275Y, showing a markedly higher frequency (32.4%) than that in other prefectures. In the 2008–09 season, however, ORVs were observed nationwide. Of 1,482 influenza viruses A (H1N1), 1,477 viruses possessed a H275Y mutation, for a total frequency of 99.7% (Figure 2, panel B). These data show that ORVs increased dramatically in Japan from the 2007–08 season to the 2008–09 season.
Genetic Analysis
Appendix Figure
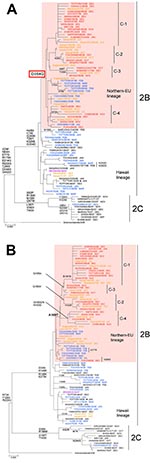
Appendix Figure. Phylogenetic analysis of influenza A (H1N1) A) neuraminidase (NA) genes and B) hemagglutinin (HA) (HA1 region) genes. Recent influenza viruses A (H1N1) fell into either clade 2B or clade 2C. Almost...
Influenza viruses A (H1N1) during 2007–08 fell into either clade 2B, including the current vaccine strain A/Brisbane/59/2007, or clade 2C, and almost all influenza viruses A (H1N1) during 2008–09 fell into clade 2B. Most ORVs with H275Y belong to clade 2B, which can be further divided into 2 distinct lineages by an aspartic acid to glycine substitution at aa 354: a Northern-Eu lineage sharing 354G, which was first isolated from countries in northern Europe and now represents most ORVs worldwide; and a Hawaii lineage sharing 354D, which was first detected in Hawaii and was rarely isolated in a few countries during the 2007–08 season (Appendix Figure). In the 2007–08 season, of 45 ORVs, 1 virus (A/Yokohama/91/2007) isolated in November 2007 belonged to clade 2C, and 44 viruses fell into either the Hawaii lineage or Northern-Eu lineage. Conversely, in the 2008–09 season, all ORVs belonged to the Northern-Eu lineage, indicating that ORVs of the Northern-Eu lineage dominated in the 2008–09 season in Japan.
In the Hawaii lineage, OSVs genetically close to ORVs were observed. The NA gene of some ORVs had only 1 nucleotide difference from that of OSV counterparts (i.e., A/Tochigi/8/2008 and A/Tochigi/9/2008, A/Nagano/1100/2008 and A/Nagano/1071/2008, A/Yamagata/68/2008 and A/Yamagata/41/2008, respectively), and the other ORVs are also genetically close to OSVs from Japan (Appendix Figure). These HA genes were also genetically identical or close together (Appendix Figure), suggesting that almost all ORVs from Japan with the Hawaii lineage are derived from OSVs from Japan. On the other hand, in the Northern-Eu lineage, OSV counterparts were not observed, but foreign ORVs genetically close to ORVs from Japan were observed. During the 2007–08 season, the NA gene of ORVs from Japan was close to that of ORVs isolated from countries in Europe (i.e., A/Paris/0341/2007 and A/England/26/2008). During the 2008–09 season, the ORVs from Japan, which shared A189T on HA protein, were further divided into 4 subclades (C-1 to C-4) by common amino acid changes on HA and/or NA (Appendix Figure). ORVs from Japan in C-2 and C-3 were genetically close to the ORVs isolated from North America or Hawaii (e.g., A/Memphis/03/2008 and A/Hawaii/19/2008), whereas ORVs in C-1, representing most influenza viruses A (H1N1) from the 2008–09 season in Japan, and ORVs in C-4 were close to ORVs isolated from South Africa and Australia in the Southern Hemisphere (e.g., A/Kenya/1432/2008 and A/Victoria/501/2008). All ORVs except C-3 were isolated before the emergence of ORVs from Japan in each subclade. These findings suggest that ORVs from Japan within a Northern-Eu lineage would not have emerged domestically but instead may have been introduced from various countries.
Antiviral Drug Susceptibility
Figure 3

Figure 3. Assessment of drug concentrations required to inhibit neuraminidase activity by 50% (IC50) for neuramindase inhibitors (NAIs). Normal sigmoid curves were generated for most tested viruses by a neuraminidase inhibition assay for...
Of the 364 viruses (306 isolates in the 2007–08 season and 58 isolates in the 2008–09 season) tested by NA inhibition assay, 101 possessed a H275Y substitution. With the NA inhibition assay, although precise IC50 values were calculated from a normal sigmoid curve (Figure 3, panels A and B), some viruses generated 2 types of unusual sigmoid curves (Figure 3, panels C and D) resulting from the mixed population of NAI-resistant and -sensitive viruses, as previously reported (18). Tentative IC50 values were calculated from type A curves (Figure 3, panel C) and included in overall statistical analysis, but values could not be calculated from type B curves (Figure 3, panel D). Later viruses were regarded as resistant candidates.
In the NA inhibition assay for oseltamivir, OSVs showed a mean IC50 ± SD of 0.10 ± 0.05 nmol/L (range 0.01–0.35 nmol/L), and ORVs had a mean ± SD IC50 of 67.7 ± 44.1 nmol/L (range 26.1–239.2 nmol/L), showing a reduction of >260-fold in susceptibility to oseltamivir. One OSV identified as a statistical outlier (cutoff IC50, >0.40 nmol/L; upper quartile + 5.0× interquartile range) showed a D151E substitution on the NA protein (Table 1).
In the NA inhibition assay for zanamivir, statistical analysis showed that 341 viruses were regarded as the zanamivir-sensitive viruses, with a mean ± SD IC50 of 0.40 ± 0.26 nmol/L (range 0.01–1.92 nmol/L), and 16 viruses (10 ORVs and 6 OSVs) were identified as outliers (cutoff IC50, >1.99 nmol/L) (Table 1). Seven viruses (2 ORVs and 5 OSVs) were regarded as resistant candidates from curve fit patterns. NA-sequencing for these 23 viruses (12 ORVs and 11 OSVs) showed 2 types of amino acid changes on the NA protein. One virus, A/Tottori/16/2008 (OSV), possessed a Q136K substitution, showing a high IC50 (41.89 nmol/L), and 19 of the other 22 viruses displayed an amino acid change G, N, or V at the D151 position (Table 1). These data suggest that D151 changes have a substantial effect on sensitivity to zanamivir (and oseltamivir). Moreover, A/Tottori/44/2008 with H275Y and D151D/G substitutions conferred high-level resistance to both NAIs (Figure 3, panels A and B). However, a recent study reported that a D151E change was detected only after virus propagation in cell culture, but not in the original clinical specimen (19), suggesting a possible role of cell culture in selecting these D151 variant viruses. To further investigate D151 variants, available original clinical specimens of viruses with D151 variation were subjected to NA sequencing, so that all D151 variations (D151G/E/N) were not detected in the original clinical specimens (Table 1). We thus concluded that D151 variants might not have emerged as a natural occurrence and all recent ORVs would retain sensitivity to zanamivir.
Susceptibility to M2 inhibitors was determined to find established-resistant markers by M2-sequencing. The 367 viruses (288 isolates in the 2007–08 season; 79 isolates in the 2008–09 season) including 123 ORVs (45 and 78, respectively) and 244 OSVs (243 and 1, respectively) were tested. Viruses belonging to clade 2B were sensitive to M2 inhibitors, and viruses belonging to clade 2C were resistant to M2 inhibitors, so all ORVs except A/Yokohama/91/2007 were sensitive to M2 inhibitors. A/Yokohama/91/2007 belonged to clade 2C and was the only virus resistant to both oseltamivir and M2 inhibitors.
Antigenic Characteristics
To estimate the reactivity of ORVs and OSVs to ferret antiserum against 2008–09 vaccine strain A/Brisbane/59/2009, the HI test was performed. Good inhibition was achieved in 76% of OSVs and 69% of ORVs by A/Brisbane/59/2009 ferret antiserum, and 22% of OSVs and 28% of ORVs showed a 4-fold reduction in HI titer to the homologous titer, respectively (Table 2). Only 2% and 3% of OSVs and ORVs showed a >8-fold reduction in HI titer to A/Brisbane/59/2009 ferret antiserum. These data demonstrated that OSVs and ORVs were anitigenitically indistinguishable from each other and were similar to the 2008–09 vaccine strain A/Brisbane/59/2009.
High Frequency of ORVs in Tottori Prefecture during the 2007–08 Season
Figure 4

Figure 4. Phylogenetic analysis of influenza A (H1N1) neuraminidase genes (A) and geographic distribution of oseltamivir-resistant viruses (ORVs) (B) isolated from Tottori Prefecture, Japan, 2007–08. ORVs fell into either Northern-Eu lineage (red) or...
Tottori Prefecture is located in the western part of the main island of Japan. Comprising 19 cities and geographically divided into 3 areas, this prefecture has the lowest population in Japan (Figure 4, panel B). Despite a low frequency of only 2.6% in Japan during 2007–08 season, an unexpectedly high frequency (32.4%) of ORVs was observed in Tottori prefecture (Figure 2, panel A). ORVs from Tottori were collected from 4 cities in 2 areas with no systematic bias apparent in the sampling process (Figure 4, panel B).
Phylogenetic analysis of NA genes showed that these ORVs formed 3 subclades (Figure 4, panel A): the first with a Northern-Eu lineage sharing V75A and D354G (T-1); the second with a Hawaii lineage without common changes (T-2); and the third with a Hawaii lineage and sharing M188L (T-3). Among these, only OSVs genetically close to ORVs were observed in T-2, suggesting that ORVs in T-2 would be derived from OSVs in Tottori prefecture.
A mapping study for ORVs showed that all ORVs in the Hawaii lineage were collected from Tottori city only, primarily at the end of January, whereas ORVs with the Northern-Eu lineage were collected from 4 cities, including Tottori city, during February and March. Genetically diverse ORVs belonging to T1-T3 were cocirculating only in Tottori city in the eastern area (Figure 4, panel B). The Tottori case raised concern about the possibility that these Tottori ORVs could survive to become an origin ORV for the 2008–09 season in Japan. However, phylogenetic analysis showed that all ORVs isolated during the 2008–09 season were not genetically close to ORVs from Tottori (Appendix Figure). As a result, all ORVs from Tottori seemed to have been eliminated in the 2007–08 season, and ORVs that may have been introduced from other counties were circulating during 2008–09 in Japan.
Our study demonstrated that ORVs dramatically increased in Japan from the 2007–08 season (2.6%) to the 2008–09 season (99.7%). All tested ORVs showed a reduction of >260-fold in susceptibility to oseltamivir by NA inhibition assay. On the other hand, almost all ORVs remained sensitive to the other antiviral-drugs, e.g., zanamivir, and M2 inhibitors. HI testing suggested that the current vaccine, A/Brisbane/59/2008, would be effective against recent ORVs. In addition, recent studies have reported that symptoms and hospitalization rates of patients infected with ORVs are no different from those seen with OSVs (14,20).
Japan has the largest per capita use of oseltamivir (>70%) in the world (10). Because this use could cause efficient selection of ORVs in individual patients, Japan might be the initial site of worldwide spread of ORVs. However, long-term NAI surveillance in Japan during 1996–2007 and recent surveillance showed a low frequency of NAI-resistant viruses for any strains and subtypes (10,21,22), suggesting that transmissibility of ORVs selected by drug pressure was remarkably decreased. In addition, previous NAI surveillance (5–10) and several animal studies (23–26) also suggested that NAI-resistant viruses would become defective viruses with attenuated infectivity and transmissibility to human. In contrast, despite little NAI use, a high emergence of ORVs has been detected in several countries in Europe since November 2007. These ORVs had as efficient transmissibility as OSVs in human-to-human transmission, resulting in worldwide spread in a short period of time. Although whether the initial ORV detected in Norway in the 2007–08 season appeared because of NAI drug pressure is unknown, those ORVs may have obtained amino acid changes on NA and/or other proteins to compensate for the defect, in addition to the H275Y substitution on the NA protein. Most ORVs belong genetically to the Northern-Eu lineage in clade 2B, suggesting that the gene constellation may contain a big advantage to retain infectivity and transmissibility.
An interesting question arose as to where the ORVs in Japan originated. In the Hawaii lineage, almost all ORVs in Japan would be derived from OSVs in Japan because the NA gene of ORVs was similar to OSV counterparts isolated at similar times or from similar regions (Appendix Figure). On the other hand, in the Northern-Eu lineage, ORVs in Japan would have been introduced from other countries. In 2007–08, almost all ORVs would be imported from countries in Europe. In 2008–09, the ORVs in C-1, which comprised most isolates in 2008–09, and ORVs in C-4 were genetically similar to ORVs isolated from the Southern Hemisphere. Because influenza activity in the Southern Hemisphere occurs half a year earlier than that in the Northern Hemisphere, most ORVs in Japan conceivably could have been imported from the Southern Hemisphere. ORVs in C-2 and C-3 were genetically similar to ORVs isolated in North America and Hawaii, but the collection month of ORVs in C-3 were similar to each other, suggesting that ORVs in C-3 might be derived from an unknown common origin ORV. The ORVs obtained during 2008–09 were not genetically similar to any ORVs isolated in Tottori during 2007–08, indicating that ORVs from Tottori had been eliminated and had not formed the origin ORVs for the 2008–09 season in Japan. As for A/Yokohama/91/2007 belonging to clade 2C, the patient from which this virus was isolated was known to have taken oseltamivir before sampling (22), indicating that selective drug pressure in this person might have selected for this ORV.
In the NA inhibition assay for zanamivir, some viruses, including ORVs and OSVs, showed reduced sensitivity to zanamivir. NA sequencing of these viruses showed 2 types of amino acid changes. One virus, A/Tottori/16/2008 (OSV), possessed a Q136K substitution, which reportedly confers resistance to zanamivir (27,28). Conversely, most of the other viruses possessed D151 G/V/N. The amino acid changes D151 to N or E among subtype H1N1 viruses and to A, G, E, N, or V among H3N2 have been reported (7,8,19), and viruses with D151 substitutions often exhibit reduced sensitivity to NAIs (8,19,29). However, a recent study reported a possible role for cell culture in selecting these D151 variant viruses (19). In the present study, D151 variations (D151G/E/N) also were not detected from available original clinical specimens (Table 1), supporting the previous finding. We thus concluded that viruses with D151 variations would not have emerged naturally, and all ORVs would remain sensitive to zanamivir.
By sequencing of M2 gene, we confirmed that almost all Japanese ORVs belonging to clade 2B retained sensitive genotype to M2 inhibitors, consistent with previously reports that recent clade 2B viruses are sensitive to M2 inhibitors, but clade 2C viruses are resistant (27).
During the 2007–09 seasons, we also addressed NAI surveillance for A/H3N2 and type B circulating in Japan and identified no viruses resistant to both NAIs. Conversely, in March and early April 2009, a new swine-origin influenza virus A (H1N1) (now known as pandemic [H1N1] 2009 virus) emerged in Mexico and the United States and spread rapidly to many countries, including Japan (30–33). In June 2009, detection of pandemic (H1N1) 2009 virus with H275Y on the NA protein was reported from Denmark, Hong Kong Special Administrative Region, People’s Republic of China, and Japan, but all ORVs of pandemic (H1N1) 2009 virus emerged as sporadic cases with no evidence of efficient human-to-human transmission (34). Although oseltamivir remains a valuable drug for treatment of pandemic (H1N1) 2009, many ORVs were isolated after prophylaxis with a half dose of the drug. Therefore, prophylaxis with oseltamivir may not be recommended as stated by WHO (35). Rapid and continuous monitoring of NAI-resistant viruses, including pandemic (H1N1) 2009 virus, and dissemination of the findings in timely manner remains essential.
Dr Ujike is a virologist and the senior researcher of Influenza Virus Research Center of the National Institute of Infectious Diseases, Tokyo, Japan. His research interests are influenza virus and severe acute respiratory syndrome–coronavirus.
Acknowledgments
We thank Miho Ejima and Mariko Tokunaga for the production of several figures.
This study was supported by Grants-in-Aid for Seisaku-Soyaku-Sougo (H18-Iyaku- Ippan 033), and Emerging and Reemerging Infectious Diseases (H20-Shinko-Ippan 005) from the Ministry of Health, Labour and Welfare of Japan.
References
- Cauchemez S, Valleron AJ, Boëlle PY, Flahault A, Ferguson NM. Estimating the impact of school closure on influenza transmission from sentinel data. Nature. 2008;452:750–4. DOIPubMedGoogle Scholar
- Fleming DM. Managing influenza: amantadine, rimantadine and beyond. Int J Clin Pract. 2001;55:189–95.PubMedGoogle Scholar
- Saito R, Li D, Suzuki H. Amantadine-resistant influenza A (H3N2) virus in Japan, 2005–2006. N Engl J Med. 2007;356:312–3. DOIPubMedGoogle Scholar
- Saito R, Suzuki Y, Li D, Zaraket H, Sato I, Masaki H, Increased incidence of adamantane-resistant influenza A(H1N1) and A(H3N2) viruses during the 2006–2007 influenza season in Japan. J Infect Dis. 2008;197:630–2. DOIPubMedGoogle Scholar
- World Health Organization. Monitoring of neuraminidase inhibitor resistance among clinical influenza virus isolates in Japan during 2003–2006 influenza seasons. Wkly Epidemiol Rec. 2007;82:149–50.PubMedGoogle Scholar
- Monto AS, McKimm-Breschkin JL, Macken C, Hampson AW, Hay A, Klimov A, Detection of influenza viruses resistant to neuraminidase inhibitors in global surveillance during the first 3 years of their use. Antimicrob Agents Chemother. 2006;50:2395–402. DOIPubMedGoogle Scholar
- McKimm-Breschkin J, Trivedi T, Hampson A, Hay A, Klimov A, Tashiro M, Neuraminidase sequence analysis and susceptibilities of influenza virus clinical isolates to zanamivir and oseltamivir. Antimicrob Agents Chemother. 2003;47:2264–72. DOIPubMedGoogle Scholar
- Sheu TG, Deyde VM, Okomo-Adhiambo M, Garten RJ, Xu X, Bright RA, Surveillance for neuraminidase inhibitor resistance among human influenza A and B viruses circulating worldwide from 2004 to 2008. Antimicrob Agents Chemother. 2008;52:3284–92. DOIPubMedGoogle Scholar
- Kramarz P, Monnet D, Nicoll A, Yilmaz C, Ciancio B. Use of oseltamivir in 12 European countries between 2002 and 2007–lack of association with the appearance of oseltamivir-resistant influenza A(H1N1) viruses. Euro Surveill. 2009;14. pii: 19112.
- Tashiro M, McKimm-Breschkin JL, Saito T, Klimov A, Macken C, Zambon M, Surveillance for neuraminidase-inhibitor-resistant influenza viruses in Japan, 1996–2007. Antivir Ther. 2009;14:751–61. DOIPubMedGoogle Scholar
- Rameix-Welti MA, Enouf V, Cuvelier F, Jeannin P, van der Werf S. Enzymatic properties of the neuraminidase of seasonal H1N1 influenza viruses provide insights for the emergence of natural resistance to oseltamivir. PLoS Pathog. 2008;4:e1000103. DOIPubMedGoogle Scholar
- Meijer A, Lackenby A, Hungnes O, Lina B, van-der-Werf S, Schweiger B, European Influenza Surveillance Scheme. Oseltamivir-resistant influenza virus A (H1N1), Europe, 2007/08 season. Emerg Infect Dis. 2009;15:552–60. DOIPubMedGoogle Scholar
- Lackenby A, Hungnes O, Dudman SG, Meijer A, Paget WJ, Hay AJ, Emergence of resistance to oseltamivir among influenza A(H1N1) viruses in Europe. Euro Surveill. 2008;13. pii: 8026.
- Hauge SH, Dudman S, Borgen K, Lackenby A, Hungnes O. Oseltamivir-resistant influenza viruses A (H1N1), Norway, 2007/08. Emerg Infect Dis. 2009;15:155–62. DOIPubMedGoogle Scholar
- World Health Organization. WHO/ECDC frequently asked questions for oseltamivir resistance. 2008 Feb 15 [cited 2009 Oct 20]. http://www.who.int/csr/disease/influenza/oseltamivir_faqs/en/index.html
- World Health Organization. Influenza A(H1N1) virus resistance to oseltamivir. 2009 Mar 21 [cited 2009 Oct 20]. http://www.who.int/csr/disease/influenza/h1n1_table/en/index.html
- World Health Organization. 2002. Manual on animal influenza diagnosis and surveillance, 2nd ed. [cited 2009 Oct 20]. http://www.wpro.who.int/NR/rdonlyres/EFD2B9A7-2265-4AD0-BC98-97937B4FA83C/0/manualonanimalaidiagnosisandsurveillance.pdf
- Wetherall NT, Trivedi T, Zeller J, Hodges-Savola C, McKimm-Breschkin JL, Zambon M, Evaluation of neuraminidase enzyme assays using different substrates to measure susceptibility of influenza virus clinical isolates to neuraminidase inhibitors: report of the neuraminidase inhibitor susceptibility network. J Clin Microbiol. 2003;41:742–50. DOIPubMedGoogle Scholar
- Deyde VM, Okomo-Adhiambo M, Sheu TG, Wallis TR, Fry A, Dharan N, Pyrosequencing as a tool to detect molecular markers of resistance to neuraminidase inhibitors in seasonal influenza A viruses. Antiviral Res. 2009;81:16–24. DOIPubMedGoogle Scholar
- Dharan NJ, Gubareva LV, Meyer JJ, Okomo-Adhiambo M, McClinton RC, Marshall SA, Infections with oseltamivir-resistant influenza A(H1N1) virus in the United States. JAMA. 2009;301:1034–41. DOIPubMedGoogle Scholar
- Tamura D, Mitamura K, Yamazaki M, Fujino M, Nirasawa M, Kimura K, Oseltamivir-resistant influenza a viruses circulating in Japan. J Clin Microbiol. 2009;47:1424–7. DOIPubMedGoogle Scholar
- Kawakami C, Obuchi M, Saikusa M, Noguchi Y, Ujike M, Odagiri T, Isolation of oseltamivir-resistant influenza A/H1N1 virus of different origins in Yokohama City, Japan, during the 2007–2008 influenza season. Jpn J Infect Dis. 2009;62:83–6.PubMedGoogle Scholar
- Ives JA, Carr JA, Mendel DB, Tai CY, Lambkin R, Kelly L, The H274Y mutation in the influenza A/H1N1 neuraminidase active site following oseltamivir phosphate treatment leave virus severely compromised both in vitro and in vivo. Antiviral Res. 2002;55:307–17. DOIPubMedGoogle Scholar
- Carr J, Ives J, Kelly L, Lambkin R, Oxford J, Mendel D, Influenza virus carrying neuraminidase with reduced sensitivity to oseltamivir carboxylate has altered properties in vitro and is compromised for infectivity and replicative ability in vivo. Antiviral Res. 2002;54:79–88. DOIPubMedGoogle Scholar
- Herlocher ML, Truscon R, Elias S, Yen HL, Roberts NA, Ohmit SE, Influenza viruses resistant to the antiviral drug oseltamivir: transmission studies in ferrets. J Infect Dis. 2004;190:1627–30. DOIPubMedGoogle Scholar
- Herlocher ML, Carr J, Ives J, Elias S, Truscon R, Roberts N, Influenza virus carrying an R292K mutation in the neuraminidase gene is not transmitted in ferrets. Antiviral Res. 2002;54:99–111. DOIPubMedGoogle Scholar
- Barr IG, Deng YM, Iannello P, Hurt AC, Komadina N. Adamantane resistance in influenza A(H1) viruses increased in 2007 in South East Asia but decreased in Australia and some other countries. Antiviral Res. 2008;80:200–5. DOIPubMedGoogle Scholar
- Hurt AC, Holien JK, Parker M, Kelso A, Barr IG. Zanamivir-resistant influenza viruses with a novel neuraminidase mutation. J Virol. 2009;83:10366–73. DOIPubMedGoogle Scholar
- Yen HL, Hoffmann E, Taylor G, Scholtissek C, Monto AS, Webster RG, Importance of neuraminidase active-site residues to the neuraminidase inhibitor resistance of influenza viruses. J Virol. 2006;80:8787–95. DOIPubMedGoogle Scholar
- Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science. 2009;325:197–201. DOIPubMedGoogle Scholar
- Novel Swine-Origin Influenza A (H1N1) Virus Investigation Team, Dawood FS, Jain S, Finelli L, Shaw MW, Lindstrom S, et al. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl J Med. 2009;360:2605–15. DOIPubMedGoogle Scholar
- Shinde V, Bridges CB, Uyeki TM, Shu B, Balish A, Xu X, Triple-reassortant swine influenza A (H1) in humans in the United States, 2005–2009. N Engl J Med. 2009;360:2616–25. DOIPubMedGoogle Scholar
- Smith GJ, Vijaykrishna D, Bahl J, Lycett SJ, Worobey M, Pybus OG, Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature. 2009;459:1122–5. DOIPubMedGoogle Scholar
- World Health Organization. Viruses resistant to oseltamivir (Tamiflu) identified. 2009 Jul 8 [cited 2009 Oct 20]. http://www.who.int/csr/disease/swineflu/notes/h1n1_antiviral_resistance_20090708/en/index.html
- World Health Organization. Antiviral use and the risk of drug resistance. Pandemic (H1N1) 2009 briefing note 12 [cited 2009 Oct 20]. http://www.who.int/csr/disease/swineflu/notes/h1n1_antiviral_use_20090925/en/index.html
Figures
Tables
Cite This Article1Members of the Working Group for Influenza Virus Surveillance in Japan: Hideki Nagano (Hokkaido Institute of Public Health), Masayuki Kikuchi (Sapporo City Institute of Public Health), Masaki Takahashi (Research Institute for Environmental Sciences and Public Health of Iwate Prefecture), Yuki Sato (Miyagi Prefectural Institute of Public Health and Environment), Masanori Katsumi (Sendai City Institute of Public Health), Hiroyuki Saito (Akita Research Center for Public Health and Environment), Katsumi Mizuta (Yamagata Prefectural Institute of Public Health), Syoko Hirose (Fukushima Prefectural Institute of Public Health), Setsuko Fukaya (Ibaraki Prefectural Institute of Public Health), Asumi Hirata(Tochigi Prefectural Institute of Public Health and Environmental Science), Hiroyuki Tsukagoshi (Gunma Prefectural Institute of Public Health and Environmental Sciences), Yuka Uno (Saitama City Institute of Health Science and Research), Hiromi Maru (Chiba Prefectural Institute of Public Health), Hajime Yokoi (Chiba City Institute of Health and Environment), Mami Nagashima (Tokyo Metropolitan Institute of Public Health), Sumi Watanabe (Kanagawa Prefectural Institute of Public Health), Hideaki Shimizu (Kawasaki City Institute of Public Health), Sumiko Ueda (Sagamihara City Laboratory of Public Health), Chika Hirokawa (Niigata Prefectural Institute of Public Health and Environmental Sciences), Gen Kobayashi and Yoko Miyajima (Niigata City Institute of Public Health and Environment), Sanae Kuramoto and Hiroe Kodama (Ishikawa Prefectural Institute of Public Health and Environmental Science), Masako Nakamura (Fukui Prefectural Institute of Public Health), Hiroyoshi Asakawa (Yamanashi Institute for Public Health), Seiko Sawatari (Gifu Prefectural Institute of Health and Enviromental Sciences), Yasunori Tanaka (Gifu Municipal Institute of Public Health), Yoshinobu Miwa (Shizuoka Institute of Environment and Hygiene), Shinobu Ide (Shizuoka City Institute of Environmental Sciences and Public Health), Yoshinori Kohno and Ryu Hibino (Hamamatsu City Health Environment Research Center), Mami Hata (Aichi Prefectural Institute of Public Health), Noriko Goto (Nagoya City Public Health Research Institute), Takuya Yano (Mie Prefecture Health and Environment Research Institute), Fumie Matsumoto (Shiga Prefectural Institute of Public Health), Tohru Ishizaki (Kyoto Prefectural Institute of Public Health and Environment), Satoshi Hiroi (Osaka Prefectural Institute of Public Health), Hideyuki Kubo (Osaka City Institute of Public Health and Environmental Sciences), Kiyoko Uchino (Sakai City Institute of Public Health), Tomohiro Oshibe (Hyogo Prefectural Institute of Public Health and Consumer Sciences), Ai Mori and Tomoko Suga (Kobe Institute of Health), Yoshiteru Kitahori (Nara Prefectural Institute for Hygiene and Environment), Fumio Terasoma (Wakayama Prefectural Research Center of Environment and Public Health), Yoshiaki Kimura and Naomi Matsumoto (Tottori Prefectural Institute of Public Health and Environmental Science), Tamaki Omura (Shimane Prefectural Institute of Public Health and Environmental Science), Shinichi Takao (Center for Public Health and Environment, Hiroshima Prefectural Technology Research Institute), Katsuhiko Abe (Hiroshima City Institute of Public Health), Yumiko Kawakami (Tokushima Prefectural Centre for Public Health and Environmental Sciences), Ootsuka Yuka (Ehime Prefecture Institute of Public Health and Environmental Science), Daisuke Kawamoto (Fukuoka City Institute for Hygiene and the Environment), Takayuki Hirano and Hisato Masumoto (Saga Prefectural Institute of Public Health and Pharmaceutical Research), Manabu Hirano and Akinori Yamaguchi (Nagasaki Prefectural Institute for Environment Research and Public Health), Seiya Harada and Koichi Nishimura (Kumamoto Prefectural Institute of Public Health and Environmental Science), Miki Kato (Oita Prefectural Institute of Health and Environment), Miho Miura (Miyazaki Prefectural Institute for Public Health and Environment), Kanji Ishitani and Akihide Kamimura (Kagoshima Prefectural Institute for Environmental Research and Pubulic Health), Katsuya Taira (Okinawa Prefectural Institute of Health and Environment), Mitsutaka Kuzuya (Okayama Prefectural Institute for Environmental Science and Public Health), Shoichi Toda (Yamaguchi Prefectural Institute of Public Health and Environment), Chiharu Kawakami (Yokohama City Institute of Health), Mayumi Konno (Kyoto City Institute of Health and Environmental Sciences), Hiroya Komoda (Kagawa Prefectural Research Institute for Environmental Sciences and Public Health), Tae Taniwaki (Kochi Public Health and Sanitation Institute), Toshitaka Minegishi (Saitama Institute of Public Health), Rika Tsutsui (Aomori Prefectural Institute of Public Health and Environment), Shizuko Kasuo (Nagano Environmental Conservation Research Institute), Yuichiro Okamura (Nagano City Health Center), Eiji Horimoto (Toyama Institute of Health), Nobuyuki Sera (Fukuoka Institute of Health and Environmental Sciences), and Hiromi Yoshikawa (Kitakyushu City Institute of Environmental Sciences)
Table of Contents – Volume 16, Number 6—June 2010
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Takato Odagiri, Influenza Virus Research Center, National Institute of Infectious Diseases, 4-7-1,Gakuen, Musashi-Murayama, Tokyo, 208-0011, Japan
Top