Volume 19, Number 12—December 2013
Dispatch
Novel Cause of Tuberculosis in Meerkats, South Africa
Cite This Article
Citation for Media
Abstract
The organism that causes tuberculosis in meerkats (Suricata suricatta) has been poorly characterized. Our genetic analysis showed it to be a novel member of the Mycobacterium tuberculosis complex and closely related to the dassie bacillus. We have named this epidemiologically and genetically unique strain M. suricattae.
Tuberculosis (TB) is caused by a group of distinct mycobacterial strains that might have evolved as host-adapted ecotypes (1) and that are collectively named the Mycobacterium tuberculosis complex (MTC) (2). In southern Africa, M. tuberculosis and M. bovis cause TB in numerous animals (3), the dassie bacillus infects rock hyraxes (dassies, Procavia capensis), and M. mungi infects banded mongooses (Mungos mungo) (4–6).
TB in free-living meerkats (Suricata suricatta) from the Kalahari Desert, South Africa, was first reported in 2002 (7), and its epidemiology and pathology have been comprehensively described (8,9). Mycobacterial strains isolated from these animals have been described as M. tuberculosis (7), M. bovis (8), and as a “member of the animal-adapted lineage of the MTC” (10), raising concerns that the occurrence of TB in these meerkats resulted from anthropogenic exposure to these pathogens and that affected meerkat populations could pose an infection risk to other wildlife, domestic animals, and humans (8). However, these studies used genetic analyses designed to differentiate between M. tuberculosis and M. bovis (7) and between these and M. africanum, M. canetti, M. microti, and M. bovis BCG (8) but not between these strains and the dassie bacillus or M. mungi. To gain greater insight into the etiology of this disease, we conducted a more comprehensive genetic analysis of mycobacterial isolates from this meerkat population.
Permission to sample meerkats was obtained from the University of Pretoria Animal Ethics Committee. Postmortem examinations were performed on 4 meerkats from the Kalahari Meerkat Project (26°58′S, 21°49′E) that had shown visible disease. Samples from lesions typical of TB in this species (8) were used to establish mycobacterial cultures in the BD BACTEC MGIT 960 Mycobacterial Detection System (Becton Dickinson, Franklin Lakes, NJ, USA) (11). Four cultures originating from 3 animals were positive by Ziehl-Neelsen stain and were grown further on Difco Middlebrook 7H10 Agar supplemented with 10% OADC Enrichment (Becton Dickinson) for 6–8 weeks, after which DNA was extracted (11). However, only 1 isolate (MK172) yielded sufficient DNA for DNA fingerprinting by the IS6110 method (12). PCRs were conducted by using either heat-killed liquid cultures or purified DNA as a template.
Isolates were screened for the presence or absence of 4 phylogenetically informative genomic regions of difference (RDs) (11), and all showed deletion of RD9 but not of RD1, RD4, and RD12. This genotype is shared by M. africanum, M. orygis, and the dassie bacillus (2,11); isolates were therefore analyzed for the presence or absence of RD1das, a genetic marker specific for the dassie bacillus (5). Because this RD was deleted in all isolates, these were subsequently analyzed for the presence or absence of N-RD25das, RD5das, and RDVirSdas (5); a G→A single-nucleotide polymorphism (SNP) in Rv1510 (Rv15101129); and a single-nucleotide deletion in Rv0911 (Rv0911389) (2). For all isolates, N-RD25das, RD5das, and RDVirSdas were deleted and Rv15101129 and Rv0911389 were present, consistent with the dassie bacillus genotype (2,5). However, although the RD5das deletion in this bacillus has been caused by the insertion of an inverted IS6110 sequence (5), for the meerkat strain, sequencing of the RD5das PCR product showed this region to be occupied by an IS6110 sequence in a forward orientation, followed by a proline-proline-glutamate gene homologue.
Spoligotyping was performed according to the internationally standardized method (13). However, we repeatedly obtained no amplification of any spacer included in this array. We investigated the possible deletion of the direct-repeat region, the genomic region analyzed by spoligotyping, by attempting to amplify by PCR selected genetic sequences upstream and downstream thereof (Table 1). This analysis confirmed that much, if not all, of the direct-repeat region had been deleted in these isolates, together with ≈3,500 bp upstream and up to 1,700 bp downstream of this region (Table 1).
Figure 1

Figure 1. . . IS6110 restriction fragment length polymorphism patterns of A) a reference strain of Mycobacterium tuberculosis (Mt14323) with selected fragment lengths indicated in kilobases, and B) M. suricattae (isolate MK172).
Additionally, genetic characterization was done by sequencing of fragments of the gyrB gene (2) and 16S rDNA (14). For all isolates, the gyrB sequence was consistent with that of M. africanum, M. pinnipedii, and the dassie bacillus (2). However, the 16S rDNA sequence differed from that of all other MTC members by having a T→G SNP at position 214 (16S rDNA214). Analysis by mycobacterial interspersed repetitive unit–variable number tandem repeats (15) identified 2 strain variants in our sample set (Table 2); IS6110 DNA fingerprint analysis (12) of isolate MK172 showed it to contain 21 copies of the IS6110 insertion sequence element (Figure 1).
Figure 2
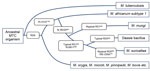
Figure 2. . . Phylogeny of the Mycobacterium tuberculosis complex (MTC) detailing relevant genetic regions of difference (RDs) and single-nucleotide differences that distinguish between M. africanum subtype 1 and the small African mammal–adapted...
We genetically characterized the causative pathogen of meerkat TB as a novel MTC strain that has several genetic features typical of the dassie bacillus and M. mungi (Figure 2). However, this pathogen differs from the closely related dassie bacillus in its mycobacterial interspersed repetitive unit–variable number tandem repeats patterns (Table 2) by being a unique RD5das variant and by containing 21 copies of the IS6110 insertion element (Figure 1) (compared with 10–15 copies in the dassie bacillus) (2). This evidence of IS6110 copy number expansion might indicate involvement of this insertion sequence in the occurrence of other genetic deletions in this strain, including those in the direct-repeat region. Notably, in addition to the novel SNP 16S rDNA214, the loss of the direct-repeat region spacers, which are routinely screened for by spoligotyping, distinguishes this strain from all other MTC members (1,13,14). As evidenced by their shared RDs and SNPs, the genetic homogeneity of multiple isolates of this distinctive strain suggests that it has undergone selective evolution, possibly through adaptation to its meerkat host (1). It is highly pathogenic in this species and seems to be substantially more virulent than the genetically similar dassie bacillus (4,5). As such, to distinguish this epidemiologically unique strain from other MTC members, we have named it M. suricattae after the host species from which it has been isolated.
The identification of this bacillus in Africa is further evidence that the early evolution of the animal-adapted MTC strains occurred on this continent. Strains derived from the early diversification of the RD9-deleted lineage include M. africanum, which has been almost exclusively isolated in West Africa (2); M. mungi, which was isolated from African mongooses (6); and M. orygis and the dassie bacillus, which have been isolated from animals mainly originating from this continent and the Middle East (2,4,5).
Of these strains, M. africanum subtype I and the dassie bacillus share a unique common progenitor (2); our study confirms the shared SNP Rv15101129 as a genetic marker thereof (Figure 2). Given that M. africanum might have an unidentified West African animal host (1), it might be useful to consider that other members of this lineage have become established in highly gregarious small mammal hosts, including 2 mongoose species.
This study demonstrates that the occurrence of TB in the Kalahari meerkats might not be indicative of an external infectious source of M. tuberculosis or M. bovis, as has been reported (6,8). Rather, our findings suggest that the disease is caused by an indigenous MTC member, which we have named M. suricattae. Our limited sample set precludes a detailed analysis of the epidemiology of this pathogen; however, the identification of this strain and the characterization of several of its discriminatory genetic markers will be useful for future investigations of the ecology and evolution of the African animal–adapted members of the MTC.
Dr Parsons is a postdoctoral fellow in the Faculty of Medicine and Health Sciences, Stellenbosch University. His research interests include the diagnosis, epidemiology, and control of TB in wildlife.
Acknowledgments
We acknowledge the expert laboratory assistance of Marianna de Kock and Claudia Spies and the logistical support of Jamie Samson. We are grateful to Tim Clutton-Brock and the Kalahari Meerkat Project for providing the meerkat carcasses.
The work was funded by the South African Medical Research Council and National Research Foundation.
References
- Smith NH, Kremer K, Inwald J, Dale J, Driscoll JR, Gordon SV, Ecotypes of the Mycobacterium tuberculosis complex. J Theor Biol. 2006;239:220–5 . DOIPubMedGoogle Scholar
- Huard RC, Fabre M, de Haas P, Lazzarini LC, van Soolingen D, Cousins D, Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex. J Bacteriol. 2006;188:4271–87. DOIPubMedGoogle Scholar
- Michel AL, Müller B, van Helden PD. Mycobacterium bovis at the animal–human interface: a problem, or not? Vet Microbiol. 2010;140:371–81. DOIPubMedGoogle Scholar
- Cousins DV, Peet RL, Gaynor WT, Williams SN, Gow BL. Tuberculosis in imported hyrax (Procavia capensis) caused by an unusual variant belonging to the Mycobacterium tuberculosis complex. Vet Microbiol. 1994;42:135–45. DOIPubMedGoogle Scholar
- Mostowy S, Cousins D, Behr MA. Genomic interrogation of the dassie bacillus reveals it as a unique RD1 mutant within the Mycobacterium tuberculosis complex. J Bacteriol. 2004;186:104–9. DOIPubMedGoogle Scholar
- Alexander KA, Laver PN, Michel AL, Williams M, van Helden PD, Warren RM, Novel Mycobacterium tuberculosis complex pathogen, M. mungi. Emerg Infect Dis. 2010;16:1296–9 . DOIPubMedGoogle Scholar
- Alexander KA, Pleydell E, Williams MC, Lane EP, Nyange JF, Michel AL. Mycobacterium tuberculosis: an emerging disease of free-ranging wildlife. Emerg Infect Dis. 2002;8:598–601 . DOIPubMedGoogle Scholar
- Drewe JA, Foote AK, Sutcliffe RL, Pearce GP. Pathology of Mycobacterium bovis infection in wild meerkats (Suricata suricatta). J Comp Pathol. 2009;140:12–24. DOIPubMedGoogle Scholar
- Drewe JA. Who infects whom? Social networks and tuberculosis transmission in wild meerkats. Proc Biol Sci. 2010;277:633–42. DOIPubMedGoogle Scholar
- Drewe JA, Eames KTD, Madden JR, Pearce GP. Integrating contact network structure into tuberculosis epidemiology in meerkats in South Africa: implications for control. Prev Vet Med. 2011;101:113–20. DOIPubMedGoogle Scholar
- Warren RM, Gey van Pittius NC, Barnard M, Hesseling A, Engelke E, de Kock M, Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference. Int J Tuberc Lung Dis. 2006;10:818–22 .PubMedGoogle Scholar
- Warren RM, van Helden PD, Gey van Pittius NC. Insertion element IS6110-based restriction fragment length polymorphism genotyping of Mycobacterium tuberculosis. Methods Mol Biol. 2009;465:353–70 .DOIPubMedGoogle Scholar
- Kamerbeek J, Schouls L, Kolk A, van Agterveld M, van Soolingen D, Kuijper S, Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology. J Clin Microbiol. 1997;35:907–14 .PubMedGoogle Scholar
- Harmsen D, Dostal S, Roth A, Niemann S, Rothgänger J, Sammeth M, RIDOM: comprehensive and public sequence database for identification of Mycobacterium species. BMC Infect Dis. 2003;3:26. DOIPubMedGoogle Scholar
- Supply P, Allix C, Lesjean S, Cardoso-Oelemann M, Rüsch-Gerdes S, Willery E, Proposal for standardization of optimized mycobacterial interspersed repetitive unit–variable-number tandem repeat typing of Mycobacterium tuberculosis. J Clin Microbiol. 2006;44:4498–510 . DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 19, Number 12—December 2013
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Sven D.C. Parsons, Division of Molecular Biology and Human Genetics, Faculty of Medicine and Health Sciences, Stellenbosch University, PO Box 19063, Tygerberg 7505, South AfricaSven D.C. Parsons, Division of Molecular Biology and Human Genetics, Faculty of Medicine and Health Sciences, Stellenbosch University, PO Box 19063, Tygerberg 7505, South Africa
Top