Volume 19, Number 9—September 2013
Research
Plasmodium falciparum Mutant Haplotype Infection during Pregnancy Associated with Reduced Birthweight, Tanzania
Cite This Article
Citation for Media
Abstract
Intermittent preventive treatment during pregnancy with sulfadoxine–pyrimethamine (IPTp-SP) is a key strategy in the control of pregnancy-associated malaria. However, this strategy is compromised by widespread drug resistance from single-nucleotide polymorphisms in the Plasmodium falciparum dihydrofolate reductase and dihydropteroate synthetase genes. During September 2008–October 2010, we monitored a cohort of 924 pregnant women in an area of Tanzania with declining malaria transmission. P. falciparum parasites were genotyped, and the effect of infecting haplotypes on birthweight was assessed. Of the genotyped parasites, 9.3%, 46.3%, and 44.4% had quadruple or less, quintuple, and sextuple mutated haplotypes, respectively. Mutant haplotypes were unrelated to SP doses. Compared with infections with the less-mutated haplotypes, infections with the sextuple haplotype mutation were associated with lower (359 g) birthweights. Continued use of the suboptimal IPTp-SP regimen should be reevaluated, and alternative strategies (e.g., intermittent screening and treatment or intermittent treatment with safe and effective alternative drugs) should be evaluated.
Pregnancy-associated malaria is a leading cause of maternal anemia and low birthweight (1). Measures to prevent pregnancy-associated malaria include insecticide-treated nets, treatment with effective antimalarial drugs, and administration of intermittent preventive treatment during pregnancy with sulfadoxine–pyrimethamine (IPTp-SP) (2). IPTp-SP is given at least twice during pregnancy, with doses 1 month apart (3,4). Studies have shown that IPTp-SP reduces the incidence of anemia, clinical malaria, low birthweight, and parasite prevalence at delivery (3,5–9).
SP acts by inhibiting the Plasmodium falciparum dihydropteroate synthetase and dihydrofolate reductase enzymes, respectively (10,11). However, resistance to the combined drug (SP) is widespread among the P. falciparum population in sub-Saharan Africa; this resistance is caused by accumulation of point mutations in the P. falciparum dihydropteroate synthetase (Pfdhfr) and dihydrofolate reductase (Pfdhps) genes (12,13). An increased number of point mutations in these genes is associated with augmented resistance to SP in vivo (14). There is sufficient evidence to support that the triple Pfdhfr mutation asparagine 51 to isoleucine (N51I), cysteine 59 to arginine (C59R), and serine 108 to asparagine (S108N) in combination with double Pfdhps mutant alanine 437 to glycine (A437G) and lysine 540 to glutamic acid (K540E)—forming quintuple mutant haplotypes—confer a high risk for treatment failure in malaria-infected children and nonpregnant adults who receive SP treatment (14). In addition, recent reports have shown that an increase in Pfdhps mutations at alanine 581 to glycine (A581G), further escalating the risk for even higher levels of resistance (15).
Because P. falciparum parasite resistance to SP is high, most likely because of the high prevalence of quintuple mutant haplotypes, use of the drug to treat uncomplicated malaria has been abandoned in many parts of eastern Africa. In Tanzania, SP was replaced in 2006 by artemether-lumefantrine for the management of uncomplicated malaria (4). The high prevalence (>50%) of the K540E mutation, which is found almost exclusively as the quintuple mutant haplotype, has also resulted in poor SP efficacy when used as intermittent preventive treatment in infants (16). However, other studies have indicated that IPTp-SP is still efficacious in some areas with high resistance (17). Nonetheless, with increased P. falciparum resistance, the usefulness of IPTp-SP might be compromised (18–20).
It has not been known whether there is an association between P. falciparum sextuple mutant haplotypes and poor pregnancy outcome. To determine if there is a relationship, we conducted a prospective cohort study in northeastern Tanzania in an area with declining malaria transmission (21). The study received ethical approval from the Tanzania Medical Research Coordinating Committee (reference no. NIMR/HQ/R.8a/Vol. IX/688). All procedures were conducted in accordance with the Declaration of Helsinki and Good Clinical and Laboratory Practices. All participants gave written informed consent.
Study Design and Samples
The study was conducted during September 2008–October 2010 in Korogwe District in the Tanga Region of northeastern Tanzania, where P. falciparum is the predominant malaria-causing species. A prospective cohort of 924 pregnant women was monitored from first attendance at the antenatal clinic through delivery. The study has been described in detail (22–24). In brief, pregnant women with a gestational age of <24 weeks, as estimated by using ultrasound, were enrolled if they had lived in Korogwe District for >6 months and were willing to give birth at Korogwe District Hospital. Study participants attended 3 additional prescheduled antenatal visits at weeks 26, 30, and 36 of pregnancy, and they were attended to by a study nurse/clinician. Obstetric history and maternal anthropometric measurements were recorded for all women (23). A venous blood sample was collected at each antenatal clinic visit, and venous and placental blood samples were collected at the time of delivery.
Diagnosis, Treatment, and Prevention of Malaria
All blood samples were tested for malaria parasites by using a rapid diagnostic test (RDT) (Parascreen, Zephyr Biomedicals, Goa, India; Paracheck Pf, Orchid Biomedical Systems, Mumbai, India; or ParaHIT-f, Span Diagnostics Ltd, Surat, India) and by microscopy. Blood smears for women with negative RDT results were examined retrospectively, whereas those for women with positive RDT results were examined immediately if deemed necessary by the study physician for a treatment decision (25). Parasite density was determined as the number of asexual stage parasites/200 leukocytes (500 leukocytes if <10 parasites) and converted to the number per microliter, as described (22,25); >100 fields were double-examined before a blood smear was declared negative.
Women with positive RDT results were treated with the antimalarial drug artemether-lumefantrine (Coartem Dispersible, Norvatis Corporation, Suffern, New York, USA) or with quinine. For infections occurring during the first trimester of pregnancy, quinine sulfate–coated tablets (ELYS Chemical Industries Ltd, Nairobi, Kenya) were used, and for severe cases, quinine dihydrochloride injection (Vital Healthcare PVT Ltd, Mumbai, India) was used.
Two doses of IPTp-SP (Sulphadar, Shelys Pharmaceutical Ltd, Dar es Salaam, Tanzania) were given >1 month apart as directly observed treatment; each dose contained 1,500 mg of sulfadoxine and 75 mg of pyrimethamine. Women with a gestational age of >20 weeks at enrollment were given the first IPTp-SP dose at the study-inclusion visit and the second dose during the third trimester. Women with a gestational age of <20 weeks at enrollment were given the first dose at 20 weeks of gestation. Women who had received IPTp-SP before study inclusion but earlier than recommended by the World Health Organization (i.e., after quickening in the second trimester) received a second dose after 20 weeks of gestation, and a third dose was given in the third trimester. Use of SP from private pharmacies/drug shops for malaria treatment before and after study inclusion was also documented. All study participants were provided with a voucher for procuring insecticide-treated nets.
Laboratory Methods and Birthweight Measurements
EDTA-preserved venous blood was used to estimate hemoglobin levels (KX-21N Automated Hematology Analyzer, Sysmex, Kobe, Japan). Live newborns whose birthweights were measured by using a spring scale (Fazzini, Vimodrone, Italy) with an accuracy of <50 g or a digital strain gauge scale (ADE, Hamburg, Germany) with an accuracy of <10 g within 24 h of delivery were included in the birthweight analysis. Newborns with severe malformations, twins, and those born to women with preeclampsia were excluded from analyses because these conditions can severely affect birthweight (23).
EDTA-preserved blood (50 μL) was spotted on Whatman number 3 filter paper (VWR– Bie & Berntsen, Herlev, Denmark), dried at room temperature, and stored in separate zip-lock bags. DNA was extracted by using the Chelex 100 method, as described (26). The DNA supernatant was transferred to a 96-well PCR plate and stored at −20°C until use. The parasite DNA was amplified by outer and nested P. falciparum–specific PCRs, as described (27); the products were analyzed by electrophoresis in 1.5% ethidium bromide–stained gel, as described (22).
To determine the multiplicity of infections, block 2 of the merozoite surface protein 2 domain was amplified by using fluorescent PCR (28). The results were analyzed by using GeneScan software, version 3.7 (Applied Biosystems, Naerum, Denmark).
Parasite DNA was amplified by outer and nested PCR with specific primers targeting the Pfdhfr and Pfdhps genes, as described (29). Single-nucleotide polymorphisms (SNPs) in the Pfdhfr and Pfdhps genes were identified by using a sequence-specific oligonucleotide probe ELISA technique, as described (29) with minor modifications. In brief, we used sequence-specific oligonucleotide probes targeting Pfdhfr codons c50/51 CI/CN, c59 (C/R), c108 (S/N/T), and c164 (I/L) and Pfdhps codons c436/437 (AA/AG/SA/SG/FG), c540 (K/E), c581 (A/G), and c613 (A/S). Individual SNPs were combined to deduce the different infecting mutant haplotypes.
Data Management and Statistical Analyses
We double-entered and validated data in Microsoft Access version 2007 (Redmond, WA, USA). Statistical analyses were conducted by using Stata version 10 (StataCorp, College Station, TX, USA) deploying parametric and nonparametric methods, as appropriate. The effect of infecting allelic haplotypes on birthweights was investigated by using multiple linear regression and dichotomized (as 6 and <6 SNPs) to infecting haplotypes; variables with a p<0.20 in univariate analysis were included in the multivariate models. By using a stepwise backward elimination approach, we obtained final models including only variables with a p<0.10. A 2-sided p-value of <0.05 was considered significant. Final models included only women without missing values.
Demographic and Parasitologic Characteristics of the Study Cohort
Of 1,171 screened pregnant women, 995 met the study inclusion criteria; 924/995 women completed follow-up. For the entire study cohort, 5,555 venous and placental blood samples were collected during antenatal care and at delivery. Among the women completing follow-up, 76 had a total of 96 episodes of malaria. Some women had >1 infection, as determined by RDT and/or blood smears. The median asexual parasite density for study participants was 2,570 asexual stages/μL (range 40–390,749).
A total of 91 samples from women with RDT parasite-positive results were examined by P. falciparum–specific PCR, and 65 (71.4%) were positive. Of these 65 samples, 54 (83.1%) from 49 women were successfully typed in subsequent molecular analyses; 21 (38.9%) of these 54 samples were obtained at study enrollment. The median age of women with samples included in the genotyping analysis was 22 years (range 17−35), and at study inclusion, they had a median gestational age of 17.7 weeks (range 6.9–23.9). Overall, 91.8% of the women in the study cohort received 2 doses of IPTp-SP.
Multiplicity of Infections and P. falciparum dhfr/dhps Genotypes
Sixty percent of the genotyped parasites were polyclonal, with a mean of 2.5 clones/sample (range 1.0–8.0 clones/sample). Of the few women with repeat P. falciparum infections and sequence-specific oligonucleotide probe ELISA data (n = 5), 1 had a recrudescent and 1 had a new infection; genotype results of the 3 remaining infections were inconclusive.
Figure 1
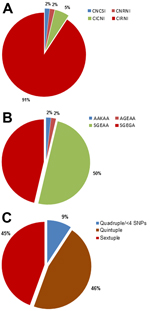
Figure 1. . . . Pfdhfr/Pfdhps haplotyping results for Plasmodium falciparum parasite isolates from 54 parasite isolates in 49 pregnant women, Korogwe District, Tanga Region, Tanzania, September 2008–October 2010. A) Proportion of single...
Of the 54 parasite isolates, 49 (90.7%) were triple Pfdhfr mutants (CIRNI [C50, 51I, 59R, 108N, and I164]); 4 (7.4%) isolates were double Pfdhfr mutants CICNI/CNRNI; and the remaining 1 (1.9%) isolates was wild-type CNCSI (Figure 1A). Two major Pfdhps haplotypes were identified in the 54 isolates: the double mutant AGEAA/SGEAA (A436/S436, A437G, and K540E) haplotype in 28 (51.9%) isolates and the triple mutant Pfdhps SGEGA (S436A, A437G, K540E, and A581G) haplotype in 25 (46.3%) isolates (Figure 1B). An AAKAA wild-type haplotype was identified in only 1 (1.9%) of the 54 isolates (Figure 1B).
The different SNPs were combined to generate the following mutant Pfdhfr/Pfdhps haplotypes: quadruple haplotype (double Pfdhfr [CICNI/CNRNI] with double Pfdhps [AGEAA/SGEAA]); quintuple haplotypes (triple Pfdhfr with double Pfdhps [CNRNI–AGEAA/SGEAA] or double Pfdhfr with triple Pfdhps [CICNI–SGEGA]); and sextuple haplotype (triple Pfdhfr [CIRNI] and triple Pfdhps [SGEGA]). The only Pfdhfr CNCSI wild-type isolate was a double Pfdhps (SGEAA) mutant, and the only Pfdhps AAKAA wild-type haplotype was a triple Pfdhfr (CIRNI) mutant. The isolates with these 2 haplotypes were grouped as quadruple or less when generating the combined Pfdhfr/Pfdhps haplotypes. The quadruple or less, quintuple, and sextuple haplotypes were observed for 9.3%, 46.3%, and 44.4% of the malaria infections, respectively (Figure 1, panel C). For the woman with repeated infections, the highest level of mutation was used in the analyses on pregnancy outcome.
Trends of Pfdhfr and Pfdhps Allelic Haplotypes by Gestational Age
Figure 2

Figure 2. . . . Proportion of mutant Plasmodium falciparum dihydrofolate reductase and dihydropteroate synthetase haplotypes among pregnant women, Korogwe District, Tanga Region, Tanzania, September 2008–October 2010. Proportions are shown by gestational age;...
Quadruple and less mutated haplotypes were observed mainly during early pregnancy, but the quintuple and sextuple haplotypes were observed throughout pregnancy (Figure 2); there was no clear trend indicating a selection by IPTp-SP use for the most mutated haplotypes (Table 1). Among the women who did not receive IPTp-SP before malaria infection, 3 with quintuple and 1 with sextuple haplotype infections reported using SP for treatment of suspected malaria before study inclusion. Even when both exposures to SP (i.e., as IPTp-SP and as SP treatment) were taken into account, no trend toward accumulation of resistant haplotypes as a result of SP use was observed (Table 1).
Effect of Sextuple Haplotypes on Pregnancy Outcome
There were no statistically significant differences in the characteristics of women infected with highly mutated parasites and those with less mutated parasites with respect to age, gravidity, and anthropometry (Table 2). Of the 26 women with quintuple or less haplotype infections and 23 with sextuple haplotype infections, 24 and 20, respectively, received 2 doses of IPTp-SP. Two women with quintuple or less haplotype infections and 2 with sextuple haplotype infections received no or 1 dose of IPTp-SP, and 1 woman with sextuple haplotype infection received 3 doses. The levels of parasitemia were also similar between the 2 groups, and there were no differences in the incidence of fever (axillary temperature >37.5°C) or level of hemoglobin (g/dL) at the time of infection or at delivery (Table 2).
Among the 49 newborns born to women for whom P. falciparum strains were genotyped, there was 1 case of preeclampsia, 2 twin deliveries, and 1 newborn with congenital malformation. Among the remaining 45 singleton newborns, 36 had their birthweight measured within 24 hours of delivery and, thus, were eligible for analysis. There was no difference in the distribution of haplotypes between mothers of the included and excluded newborns (data not shown).
The median birthweight for newborns of women with sextuple haplotype infections was significantly lower than that for newborns of women with less mutated haplotype infections (Table 2). When using z-scores (adjusting for gestational age at delivery and sex of newborn) based on a Tanzanian reference chart from the same cohort (23), borderline significance toward lower z-scores in women with sextuple mutant haplotype infections was observed (p = 0.06). There was no difference in the incidence of low birthweight newborns or of newborns that were small for gestational age at birth among women with sextuple haplotype and less mutated haplotype infections (Table 2). When the effect on birthweight was further analyzed by using a stepwise multiple linear regression model adjusting for covariates, newborns of women with sextuple mutant haplotype infections had a significantly reduced birthweight (mean reduction 359 g; 95% CI, −601 to −117) compared with newborns of women with quintuple or less haplotype infections (p = 0.005) (Table 3) Covariates for the linear regression model were mid upper arm circumference at study inclusion, parasite density, hemoglobin level, number of IPTp-SP doses, gestational age at inclusion and at delivery, number of infections, maternal age, weight/weight gain during pregnancy, HIV status, place of delivery, sex of the newborn, and gravidity.
Women in whom sextuple haplotype parasites were identified more often had repeated infections than those in whom less mutated haplotypes were identified (Table 2). In the univariate analysis, there was a borderline significance toward decreased birthweight after repeated infections (p = 0.08). However, this effect was not observed in the multivariate model (Table 3).
There was a trend toward decreased birthweight with increasing numbers of IPTp-SP doses received (Table 3). This trend was explained by the fact that newborns of the 3 women who received 0 or 1 IPTp-SP dose had much higher birthweights than newborns of the 33 women who received 2 doses (3,690 g vs. 2,920 g, respectively; p = 0.04). However, in the entire cohort (924 women) there was no association between the number of IPTp-SP doses and birthweight (data not shown). Furthermore, the number of IPTp-SP doses received before time of infection and the time between the most recent IPTp-SP dose and when infection occurred was not associated with birthweight. Adding these variables to the multivariate model did not alter the result (data not shown).
IPTp-SP is still being used in areas where prevalence of Pfdhfr/Pfdhps mutations is high (17), although widespread resistance is likely to affect the protective effect of this strategy in preventing pregnancy-associated malaria. The continued use of this suboptimal drug for IPTp might promote further development of resistance because resistant strains are likely to have a fitness advantage.
This study showed that despite administration of a full IPTp-SP course to most of the study participants, the women were not fully protected, and malaria infections occurred in 8.2% (76/924) of the women. Point mutations in Pfdhfr and/or Pfdhps genes were observed in all genotyped parasites. Quadruple or less mutated haplotypes were mainly observed early during pregnancy, whereas quintuple and sextuple mutated haplotypes were encountered throughout pregnancy. Harrington et al. (20) indicated that IPTp-SP use was associated with increased prevalence of parasites with mutations at codon A581G. However, we did not observe an association between exposure to SP before infection and increased A581G mutant parasite prevalence (Table 1). This lack of association in this study could be due to the fact that the parasite populations were already largely saturated with the highly mutated strains.
Of concern, the high prevalence of the Pfdhfr/Pfdhps sextuple haplotype was associated with reduced birthweight. Previous studies have reported an association between mutation level and adverse pregnancy outcomes (18–20) rather than, as we show, an adverse effect on birthweight specifically associated with sextuple haplotype parasites. For the entire cohort of 924 women, a negative association between malaria and fetal growth/birthweight has been reported (25).
We converted birthweights to z-scores by using a reference chart of birthweights for infants born to healthy women in the 924-person study cohort (23). By using the z-scores, we standardized the difference between individual birthweight and the mean population birthweight adjusted for sex of newborn and gestational age at delivery. The disadvantage of this conversion is that any inherent uncertainty in the reference chart is imputed into the birthweight measure. This could explain why the difference in z-scores between the sextuple and less mutated haplotypes was only borderline significant (Table 2). The multivariate model on the effect of infection with the sextuple haplotype on birthweight was already adjusted for gestational age at delivery, and the results were not altered when sex of the newborn was included (data not shown).
Harrington et al. (18) observed that the use of IPTp-SP increased parasite growth. We did not find a substantial difference in parasite density between the different haplotypes, nor did we find that the time between IPTp-SP use and infection altered the effect of the haplotype on birthweight. Therefore, IPTp-SP does not seem to increase parasite density, and differences in parasite density cannot explain the observed effect of the sextuple haplotype on birthweight. The presence of highly mutated P. falciparum genotypes that cannot be cleared by IPTp-SP exposes women to persistent and chronic malaria infections. In our study, sextuple haplotype parasite infections were associated with reduced weight of the placenta, although this difference did not reach statistical significance. Low placental weight indicates poor development of the placenta, which has been associated with pregnancy-associated malaria (30, 31). In vitro assays with extravillous trophoblasts showed that serum and plasma from P. falciparum–infected pregnant women inhibited extravillous trophoblast invasion and migration, offering a possible explanation for the pathophysiologic events that may cause impaired placentation, reduced placental weight, and low infant birthweight (32). Therefore, longer lasting infections, altering placental development among the women with sextuple haplotype infections, could explain the reduced birthweights observed in our study.
Harrington et al. (18) reported that SP in itself could have detrimental effects on the health of newborns. Our findings do not indicate that SP in itself causes lower birthweight. We did find a borderline significant association between the number of IPTp-SP doses and reduced birthweight among the 36 newborns; however, this association was not seen when we evaluated the effect of IPTp-SP doses on birthweight in the entire cohort of 924 women (data not shown).
Despite regular screening, using RDTs, for malaria among pregnant women and treatment with effective antimalarial drugs, it is worrisome that birthweights for newborns of women infected with sextuple mutated parasites were lower than those for newborns of women infected with less mutated parasites. This finding underscores the need to evaluate the effect of these mutations among populations using IPTp-SP at a wider scale.
Because of the changing epidemiology of malaria, which has transformed the transmission pattern from high to low in large parts of sub-Saharan Africa (21,33), including the study area, it is likely that many pregnant women, irrespective of gravidity, will have little immunity against pregnancy-associated malaria. The decreasing prevalence of malaria and the fact that isolates with sextuple mutations had a significant effect on birthweight underscores the question of whether IPTp-SP should be continued or replaced by intermittent screening and treatment and/or with an alternative drug for IPTp. SP might be teratogenic in the first trimester of pregnancy (34), and most other currently available antimalarial drugs are either teratogenic or their efficacy and safety profiles among pregnant women are still poorly known (35); thus, there is an urgent need to conduct further efficacy and safety studies to determine alternative drugs for IPTp. Until then, we suggest that screening by RDTs during pregnancy may allow early case detection and prompt treatment with effective antimalarial drugs (22).
This study demonstrates that sextuple Pfdhfr/Pfdhps mutated haplotypes are prevalent in the study area and that these highly SP-resistant parasites are associated with a significant reduction in birthweight of newborns of malaria-infected women. Presumably, the presence of these highly mutated P. falciparum genotypes is largely unaffected by IPTp-SP and expose women to persistent and chronic malaria infection, and this effect, rather than SP by itself, has detrimental effects on the health of newborns. We observed highly mutated haplotypes even before IPTp-SP was used and throughout pregnancy, indicating saturation of the population with resistant parasites. Therefore, continued use of the suboptimal IPTp-SP regimen should urgently be reevaluated, and its replacement with screening and treatment or with an alternative safe and effective antimalarial drug for IPTp should be considered.
Mr Minja is a research scientist at the National Institute for Medical Research, Tanzania. He is a PhD student with a keen interest in the molecular epidemiology and pathophysiology of pregnancy-associated malaria.
Acknowledgments
We thank all study participants for their willingness to participate; Tilaus Gustav, Thomson Mwampamba, Rashid Madebe, Deusdedith Makingi, Deusdedith Ishengoma, and Ulla Abildtrup for excellent technical support; Charles Tunuka, Sophia Kabome, Lydia Massawe, Francis Assenga, Latifa Shaweji, Prisca Mavindi, Rose Mutua, Halima Mpambile, and Aziz Seiph for clinical support; Christopher Mhagama, Hassan Kilavo, Silas Msangi, Stella Mkandawile, and Eva Rimoy for timely and organized data management; and Allen Mrango, Francis Mkongo, and Frank Mnango for excellent logistics support; Hannah Elena Suhrs, Martyna Gassowski, Pernille Kofoed, Line Holm, Peter Cordes, Alisha Walker, Neeltje Rutten, Nicolien Beld, and the staff of National Institute for Medical Research–Korogwe Research Laboratory for their wonderful support and cooperation; Phillip Deloron for assisting in merozoite surface protein 2 genotyping and overall coordination; Christopher Masaka, Obedi Ole Kaondo, Lydia Lugomora, and Bibiana Reuben for administrative support; and the National Institute for Medical Research–Tanga Centre and Headquarters’ Administration and Joint Malaria Programme Office and Korogwe District Hospital for their wonderful administrative support. Vito Baraka is also thanked for his useful comments.
This work was supported by the European Union through the Seventh Framework Programme STOPPAM (contract no. 200889) and the Danish International Development Agency (grant no. DFC file no.87-08-KU).
D.T.R.M., C.S., A.S., A.J.F.L., M.L., T.T., J.L., and M.A. designed the study. D.T.R.M., C.S., S.B., M.O., P.M., C.P., D.J., and J.L. conducted the study and participated in the laboratory analyses. D.T.R.M. drafted the manuscript. C.S., B.M., MA, and T.T. assisted in data management and analyses.
References
- Desai M, ter Kuile FO, Nosten F, McGready R, Asamoa K, Brabin B, Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis. 2007;7:93–104. DOIPubMedGoogle Scholar
- World Health Organization. Regional Office for Africa, Brazzaville, 2004. A strategic framework for malaria prevention and control during pregnancy in the African region. 2004 [cited 2013 Jun 12]. http://whqlibdoc.who.int/afro/2004/AFR_MAL_04.01.pdf
- Greenwood B. The use of anti-malarial drugs to prevent malaria in the population of malaria-endemic areas. Am J Trop Med Hyg. 2004;70:1–7 .PubMedGoogle Scholar
- Ministry of Health and Social Welfare, United Republic of Tanzania. National guidelines for malaria diagnosis and treatment. 2006 [cited 2013 Jun 12]. http://apps.who.int/medicinedocs/en/m/abstract/Js19271en/
- Ramharter M, Schuster K, Bouyou-Akotet MK, Adegnika AA, Schmits K, Mombo-Ngoma G, Malaria in pregnancy before and after the implementation of a national IPTp program in Gabon. Am J Trop Med Hyg. 2007;77:418–22 .PubMedGoogle Scholar
- Rogerson SJ, Chaluluka E, Kanjala M, Mkundika P, Mhango C, Molyneux ME. Intermittent sulfadoxine-pyrimethamine in pregnancy: effectiveness against malaria morbidity in Blantyre, Malawi, in 1997–99. Trans R Soc Trop Med Hyg. 2000;94:549–53. DOIPubMedGoogle Scholar
- Shulman CE, Dorman EK, Cutts F, Kawuondo K, Bulmer JN, Peshu N, Intermittent sulphadoxine-pyrimethamine to prevent severe anaemia secondary to malaria in pregnancy: a randomised placebo-controlled trial. Lancet. 1999;353:632–6. DOIPubMedGoogle Scholar
- ter Kuile FO, van Eijk AM, Filler SJ. Effect of sulphadoxine-pyrimethamine resistance on the efficacy of intermittent preventive therapy for malaria control during pregnancy: a systematic review. JAMA. 2007;297:2603–16. DOIPubMedGoogle Scholar
- van Eijk AM, Ayisi JG, ter Kuile FO, Otieno JA, Misore AO, Odondi JO, Effectiveness of intermittent preventive treatment with sulphadoxine-pyrimethamine for control of malaria in pregnancy in western Kenya: a hospital-based study. Trop Med Int Health. 2004;9:351–60. DOIPubMedGoogle Scholar
- Bzik DJ, Li WB, Horii T, Inselburg J. Molecular cloning and sequence analysis of the Plasmodium falciparum dihydrofolate reductase-thymidylate synthase gene. Proc Natl Acad Sci U S A. 1987;84:8360–4. DOIPubMedGoogle Scholar
- Sibley CH, Hyde JE, Sims PFG, Plowe CV, Kublin JG, Mberu EK, Pyrimethamine-sulfadoxine resistance in Plasmodium falciparum: what next? Trends Parasitol. 2001;17:582–8. DOIPubMedGoogle Scholar
- Hyde JE. Point mutations and pyrimethamine resistance in Plasmodium falciparum. Parasitol Today. 1989;5:252–5. DOIPubMedGoogle Scholar
- Kyabayinze D, Cattamanchi A, Mamya MR, Rosenthal PJ, Dorsey G. validation of a simplified method for using molecular markers to predict sulfadoxine-pyrimethamine treatment failure in African children with falciparum malaria. Am J Trop Med Hyg. 2003;69:247–52 .PubMedGoogle Scholar
- Kublin JG, Dzinjalamala FK, Kamwendo DD, Malkin EM, Cortese JF, Martino LM, Molecular markers for failure of sulfadoxine-pyrimethamine and chlorproguanil-dapsone treatment of Plasmodium falciparum malaria. J Infect Dis. 2002;185:380–8. DOIPubMedGoogle Scholar
- Gesase S, Gosling RD, Hashim R, Ord R, Naidoo I, Madebe R, High resistance of Plasmodium falciparum to sulphadoxine/pyrimethamine in northern Tanzania and the emergence of dhps resistance mutation at codon 581. PLoS ONE. 2009;4:e4569. DOIPubMedGoogle Scholar
- World Health Organization. WHO policy recommendation on intermittent preventive treatment during infancy with sulphadoxine-pyrimethamine (SP-IPTi) for Plasmodium falciparum malaria control in Africa. 2010 [cited 2013 Jun 12]. http://www.who.int/malaria/news/WHO_policy_recommendation_IPTi_032010.pdf
- World Health Organization; WHO Evidence Review Group. Intermittent preventive treatment of malaria in pregnancy (IPTp) with sulfadoxine-pyrimethamine (SP). 2012 [cited 2013 Jun 12]. http://www.who.int/malaria/mpac/sep2012/iptp_sp_erg_meeting_report_july2012.pdf
- Harrington WE, Morrison R, Fried M, Duffy PE. Intermittent preventive treatment in pregnant women is associated with increased risk of severe malaria in their offspring. PLoS ONE. 2013;8:e56183. DOIPubMedGoogle Scholar
- Harrington WE, Mutabingwa TK, Kabyemela E, Fried M, Duffy PE. Intermittent treatment to prevent pregnancy malaria does not confer benefit in an area of widespread drug resistance. Clin Infect Dis. 2011;53:224–30. DOIPubMedGoogle Scholar
- Harrington WE, Mutabingwa TK, Muehlenbachs A, Sorensen B, Bolla MC, Fried M, Competitive facilitation of drug-resistant Plasmodium falciparum malaria parasites in pregnant women who receive preventive treatment. Proc Natl Acad Sci U S A. 2009;106:9027–32. DOIPubMedGoogle Scholar
- Mmbando BP, Vestergaard L, Kitua A, Lemnge M, Theander T, Lusingu J. A progressive declining in the burden of malaria in north-eastern Tanzania. Malar J. 2010;9:216. DOIPubMedGoogle Scholar
- Minja DT, Schmiegelow C, Oesterholt M, Magistrado P, Bostrom S, John D, Reliability of rapid diagnostic tests in diagnosing pregnancy-associated malaria in north-eastern Tanzania. Malar J. 2012;11:211. DOIPubMedGoogle Scholar
- Schmiegelow C, Scheike T, Oesterholt M, Minja D, Pehrson C, Magistrado P, Development of a fetal weight chart using serial trans-abdominal ultrasound in an East African population: a longitudinal observational study. PLoS ONE. 2012;7:e44773. DOIPubMedGoogle Scholar
- Schmiegelow C, Minja D, Oesterholt M, Pehrson C, Suhrs H, Boström S, Factors associated with and causes of perinatal mortality in northeastern Tanzania. Acta Obstet Gynecol Scand. 2012;91:1061–8. DOIPubMedGoogle Scholar
- Schmiegelow C, Minja D, Oesterholt M, Pehrson C, Suhrs HE, Boström S, Malaria and fetal growth alterations in the 3rd trimester of pregnancy: a longitudinal ultrasound study. PLoS ONE. 2013;8:e53794. DOIPubMedGoogle Scholar
- Wooden J, Kyes S, Sibley CH. PCR and strain identification in Plasmodium falciparum. Parasitol Today. 1993;9:303–5. DOIPubMedGoogle Scholar
- Snounou G, Viriyakosol S, Xin PZ, Jarra W, Pinheiro L, do Rosario VE, High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol. 1993;61:315–20. DOIPubMedGoogle Scholar
- Jafari S, Bras JL, Bouchaud O, Durand R. Plasmodium falciparum clonal population dynamics during malaria treatment. J Infect Dis. 2004;189:195–203. DOIPubMedGoogle Scholar
- Alifrangis M, Enosse S, Pearce R, Drakeley C, Roper C, Khalil IF, A simple, high-throughput method to detect Plasmodium falciparum single nucleotide polymorphisms in the dihydrofolate reductase, dihydropteroate synthase, and P. falciparum chloroquine resistance transporter genes using polymerase chain reaction– and enzyme-linked immunosorbent assay–based technology. Am J Trop Med Hyg. 2005;72:155–62 .PubMedGoogle Scholar
- Adebami OJ, Owa JA, Oyedeji GA, Oyelami OA, Omoniyi-Esan GO. Associations between placental and cord blood malaria infection and fetal malnutrition in an area of malaria holoendemicity. Am J Trop Med Hyg. 2007;77:209–13 .PubMedGoogle Scholar
- Griffin JB, Lokomba V, Landis S, Thorp J, Herring A, Tshefu A, Plasmodium falciparum parasitaemia in the first half of pregnancy, uterine and umbilical artery blood flow, and foetal growth: a longitudinal Doppler ultrasound study. Malar J. 2012;11:319. DOIPubMedGoogle Scholar
- Umbers AJ, Stanisic DI, Ome M, Wangnapi R, Hanieh S, Unger HW, Does malaria affect placental development? Evidence from in vitro models. PLoS ONE. 2013;8:e55269. DOIPubMedGoogle Scholar
- World Health Organization. World malaria report 2012 [cited 2013 Jun 12]. http://www.who.int/malaria/publications/world_malaria_report_2012/wmr2012_full_report.pdf
- Peters PJ, Thigpen MC, Parise ME, Newman RD. Safety and toxicity of sulfadoxine/pyrimethamine: implications for malaria prevention in pregnancy using intermittent preventive treatment. Drug Saf. 2007;30:481–501 . DOIPubMedGoogle Scholar
- Manyando C, Kayentao K, D'Alessandro U, Okafor H, Juma E, Hamed K. A systematic review of the safety and efficacy of artemether-lumefantrine against uncomplicated Plasmodium falciparum malaria during pregnancy. Malar J. 2012;11:141. DOIPubMedGoogle Scholar
Figures
Tables
Cite This Article1Current affiliation: Mère et Enfant Face aux Infections Tropicales, Paris, France.
Table of Contents – Volume 19, Number 9—September 2013
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Daniel T.R. Minja, National Institute for Medical Research, Tanzania–Laboratory Sciences, PO Box 5004, Tanga, United Republic of Tanzania
Top