Volume 20, Number 8—August 2014
Letter
Sika Deer Carrying Babesia Parasites Closely Related to B. divergens, Japan
Cite This Article
Citation for Media
To the Editor: Human babesiosis caused by Babesia divergens, a zoonotic pathogen of bovines in Europe, is an emerging tickborne disease (1). In the United States, a closely related Babesia sp. was identified in persons in Missouri and Kentucky and in eastern cottontail rabbits (Sylvilagus floridanus) on Nantucket Island, Massachusetts (2–5). We report that sika deer (Cervus nippon) in Japan also carry parasites genetically closely related to B. divergens.
During November 2007–February 2008 and November 2008–February 2009, we collected blood samples from 96 wild sika deer throughout Japan. We purified DNA from the blood, which had been stored in a freezer, by phenol/chloroform extraction and performed nested PCR for B. divergens 18S rRNA (rDNA), chaperonin-containing T-complex protein 1 eta subunit (CCT7, formerly described as CCTη) (6), and β-tubulin (7) genes. Primers for rDNA were designed from the sequences of related Babesia spp. (GenBank accession nos. U16370, U16369, and AY046575): dv101F (5′-ACAACAGTTATAGTTTCTTTGGTATTCG-3′) and dv1353R (5′-GCCTTAAACTTCCTTGCGGCTTAGAGC-3′), and dv159F (5′-GCTAATGCAAGTTCGAGGCCTTTTGGCG-3′) and dv1296R (5′-CGGACGAACCTTTTTACGGACACTAG-3′) for the first and second rounds, respectively. CCT7 primers were similarly designed (GenBank accession nos. AB367924 and AB367925): Bdiv/odoCCTF1 (5′-CAAAATGAGYCACCTMCTCAACCTACC-3′) and BdivCCTR1 (5′-ATCTCAGCAGCTCACTACAGTGACCACCTC-3′), and Bdiv/odoCCTF2 (5′- CAACCTACCRATTCTCCTYYTGAAGGAGGG-3′) and BdivCCTR2 (5′-GGCTAATAAGTCGATATTGCGGGGCTCACG-3′) for the first and second rounds, respectively. The β-tubulin PCR protocol has been described (7).
Of the 96 blood samples, 12 from 5 prefectures (Hokkaido, Iwate, Tochigi, Nagano, and Miyazaki) were positive for Babesia rDNA (Technical Appendix). The sequence for sample 08–22 from Hokkaido (GenBank accession no. KC465978) was distinct from sequences of the other 11 samples (97.5%–97.6% identity in 1,041 bp), which consisted of 7 variant sequences (GenBank accession nos. KC465973–7 and AB857845–6) and 5 identical sequences (GenBank accession nos. KC465977 and AB861504–7) (99.7%–100% identity). The 5 identical sequences varied in only 1 of 909 bp from B. divergens rDNA from an Ixodes persulcatus tick in Russia [GenBank accession no. GU057385] (8).
β-tubulin (900 bp) was also amplified from the 12 Babesia rDNA–positive samples. Ten of the sequences consisted of 3 sequence variants (99.9% identity; GenBank accession nos. KC465969, KC465970, and KC465968/AB861508–14). The 2 divergent sequences (GenBank accession nos. KC465971 [08–22] and KC465989 [08–25]) were most similar to B. odocoilei (GenBank accession no. KC465972; 91% identity) and Theileria orientalis (GenBank accession no. AP011947; 79.9% identity), respectively. Thus, at least 1 deer likely had Babesia and Theileria spp. infections.
CCT7 was amplified from 10 of the 12 Babesia-positive blood samples. The sequences (GenBank accession nos. KC465979–88) were more heterogeneous (98.7%–99.9% identity) than those for rDNA and β-tubulin; this finding was expected because CCT7 evolves more quickly (6).
Figure
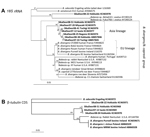
Figure. Neighbor-joining phylogenetic trees generated from Babesia sequences from GenBank and from our study in Japan, November 2007–February 2008 and November 2008–February 2009Bootstrap support (1,000 repetitions) is indicated at the nodesThe trees...
We generated a neighbor-joining phylogenetic tree (ClustalW, http://clustalw.ddbj.nig.ac.jp/index.php?lang=ja) from the Babesia rDNA sequences from our study and from GenBank (Figure, panel A). The tree shows a distinct lineage (Asian) cluster for the deer parasites, except for 08–22 (GenBank accession no. KC465978), within a clade also holding the B. divergens strains (human and bovine) from Europe (European Union lineage). The tree also shows a cluster encompassing Babesia spp. (human and rabbit) from the United States; B. divergens (deer), B. capreoli (deer), and Babesia sp. (chamois) from Europe; and B. divergens (human) from Portugal. Sequence 08–22 branches with Babesia spp. in Ix. ovatus ticks from Japan (GenBank accession nos. AY190123 and AY190124) (9). The branch lengths indicate clear separation between the isolates from sika deer and ticks, suggesting that the clustering may be attributable to the limited number of available related sequences.
We also generated a phylogenetic tree of β-tubulin sequences (900 bp), which produced similar topology and high bootstrap support (Figure, panel B). However, the limited number of relevant β-tubulin sequences precludes conclusions regarding the phylogeny of Babesia parasites. A CCT7 phylogenetic tree was not generated because of the paucity of sequences. Overall, the phylogenies suggest that B. divergens– and B. capreoli–related parasites are found worldwide in temperate zones of the Northern Hemisphere, including Europe (1), the United States (2–5), Russia (8), and Japan.
We showed the presence of B. divergens–like rDNA, β-tubulin, and CCT7 genes in sika deer from different Japanese prefectures, confirming the presence of this parasite in Japan. B. capreoli, which is serologically indistinguishable from B. divergens, was previously reported in sika deer (10). However, no molecular data for the B. capreoli isolate exist, so no conclusion may be drawn regarding its relationship to the B. divergens–like parasites from sika deer in our study.
There is an overabundance of wild sika deer in Japan because these animals easily adapt to a variety of climates, vegetation, and geography. Increased human exposure to deer habitats increases the risk of exposure to tickborne zoonoses, such as those caused by Babesia spp. In humans, infections caused by B. divergens and B. divergens–like parasites can be life threatening; fatality rates of 42% and 33% have been reported in infected asplenic patients in Europe and the United States, respectively (1–3). The findings from our study emphasize the need for increased clinical awareness of babesiosis in Japan and globally. They also emphasize the need for the swift diagnosis of suspected cases and prompt treatment of confirmed cases, especially in asplenic patients at high risk for the potentially deadly consequences of babesiosis.
Acknowledgments
We are grateful to Danny H.-K. Jokl for his critical reading of the manuscript. We thank Jeremy Gray for providing genomic DNA of B. divergens MRNK and Marek Sawczuk and Bogumiła Skotarczak for providing genomic DNA of B. divergens from Ix. ricinus tick collected in Poland.
Financial support for this study was provided in part by grants-in-aid from the Ministry of Education, Science and Culture of Japan and by the Ministry of Health, Labor and Welfare Science of Japan (H25-Shinko-Ippan-008 and H25-Shinko-Shitei-009). Support was also provided by a Grant-in-Aid for Cooperative Research from Rakuno-Gakuen University and by Gakujutsu Frontier Cooperative Research in Rakuno-Gakuen University.
References
- Gorenflot A, Moubri K, Precigout E, Carcy B, Schetters TP. Human babesiosis. Ann Trop Med Parasitol. 1998;92:489–501 . DOIPubMedGoogle Scholar
- Herwaldt B, Persing DH, Precigout EA, Goff WL, Mathiesen DA, Taylor PW, A fatal case of babesiosis in Missouri: identification of another piroplasm that infects humans. Ann Intern Med. 1996;124:643–50. DOIPubMedGoogle Scholar
- Beattie JF, Michelson ML, Holman PJ. Acute babesiosis caused by Babesia divergens in a resident of Kentucky. N Engl J Med. 2002;347:697–8. DOIPubMedGoogle Scholar
- Goethert HK, Telford SR III. Enzootic transmission of Babesia divergens among cottontail rabbits on Nantucket Island, Massachusetts. Am J Trop Med Hyg. 2003;69:455–60 .PubMedGoogle Scholar
- Spencer AM, Goethert HK, Telford SR III, Holman PJ. In vitro host erythrocyte specificity and differential morphology of Babesia divergens and a zoonotic Babesia sp. from eastern cottontail rabbits (Sylvilagus floridanus). J Parasitol. 2006;92:333–40. DOIPubMedGoogle Scholar
- Nakajima R, Tsuji M, Oda K, Zamoto-Niikura A, Wei Q, Kawabuchi-Kurata T, Babesia microti–group parasites compared phylogenetically by complete sequencing of the CCTeta gene in 36 isolates. J Vet Med Sci. 2009;71:55–68. DOIPubMedGoogle Scholar
- Hirata H, Kawai S, Maeda M, Jinnai M, Fujisawa K, Katakai Y, Identification and phylogenetic analysis of Japanese macaque Babesia-1 (JM-1) detected from a Japanese macaque (Macaca fuscata fuscata). Am J Trop Med Hyg. 2011;85:635–8. DOIPubMedGoogle Scholar
- Rar VA, Epikhina TI, Livanova NN, Panov VV. Genetic diversity of Babesia in Ixodes persulcatus and small mammals from North Ural and West Siberia, Russia. Parasitology. 2011;138:175–82 and. DOIPubMedGoogle Scholar
- Inokuma H, Yoshizaki Y, Shimada Y, Sakata Y, Okuda M, Onishi T. Epidemiological survey of Babesia species in Japan performed with specimens from ticks collected from dogs and detection of new Babesia DNA closely related to Babesia odocoilei and Babesia divergens DNA. J Clin Microbiol. 2003;41:3494–8. DOIPubMedGoogle Scholar
- Gray JS, Murphy TM, Waldrup KA, Wagner GG, Blewett DA, Harrington R. Comparative studies of Babesia spp. from white-tailed and sika deer. J Wildl Dis. 1991;27:86–91. DOIPubMedGoogle Scholar
Figure
Cite This Article1Deceased.
Related Links
Table of Contents – Volume 20, Number 8—August 2014
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Aya Zamoto-Niikura, Division of Experimental Animal Research, National Institute of Infectious Diseases, 4-7-1 Gakuen, Musashimurayama, Tokyo 208-0011, Japan
Top