Volume 21, Number 12—December 2015
Dispatch
Vectorborne Transmission of Leishmania infantum from Hounds, United States
Cite This Article
Citation for Media
Abstract
Leishmaniasis is a zoonotic disease caused by predominantly vectorborne Leishmania spp. In the United States, canine visceral leishmaniasis is common among hounds, and L. infantum vertical transmission among hounds has been confirmed. We found that L. infantum from hounds remains infective in sandflies, underscoring the risk for human exposure by vectorborne transmission.
Leishmaniasis is endemic to 98 countries (1). Canids are the reservoir for zoonotic human visceral leishmaniasis (VL) (2), and canine VL was detected in the United States in 1980 (3). Subsequent investigation demonstrated that many US hounds were infected with Leishmania infantum (4). Evidence has demonstrated that L. infantum was spread by vertical transmission over many canine generations; no evidence of vector transmission has been reported (5,6). Vertical transmission may lead L. infantum to adapt to vectorless transmission and shed largely unrecognized factors needed for vector infection. Continuous axenic cell culture conditions without vector involvement have been shown to attenuate pathogen infectivity (7). Similarly, L. infantum circulating primarily via vertical transmission within US hunting hounds may lose its ability to infect and may be transmitted by traditional vectors.
In North America, 3 species of sandfly (Lutzomyia anthophora, Lu. diabolica, and Lu. shannoni) are known vectors of Leishmania spp. Reported cases of autochthonous cutaneous leishmaniasis in the United States include 9 cases in northeastern Texas (8), 2 in Oklahoma (9), and 1 in North Dakota (10). In the Americas, the principal sandfly vector is Lu. longipalpis, which can transmit Leishmania of multiple species. (11); its northernmost distribution is limited to Mexico. Lu. shannoni sandflies have been found in Kansas and Missouri (total range 21 states) (12). During 2010–2013, we assessed whether L. infantum circulating among hunting dogs in the United States can fully develop within sandflies and be transmitted to a susceptible vertebrate host.
Figure 1
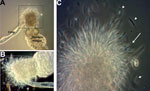
Figure 1. Sandflies infected with Leishmania infantum from US foxhounds, showing blocked stomodeal valve. Development of L. infantum (MCAN/US/2001/ FOXYMO1) in laboratory-reared Lutzomyia longipalpis sandflies led to stomodeal valve blockage 10–13 days after...
A total of 300 laboratory-reared female Lu. longipalpis sandflies were allowed to feed on 2 hounds naturally infected with L. infantum, strain MCAN/US/2001/FOXYMO1 or a closely related strain. During 2007–2011, the hounds had been tested for infection with Leishmania spp. by ELISA, PCR, and Dual Path Platform Test (Chembio Diagnostic Systems, Inc. Medford, NY, USA (Table 1). L. infantum development in these sandflies was assessed by dissecting flies starting at 72 hours after feeding and every other day thereafter. Migration and attachment of parasites to the stomodeal valve of the sandfly and formation of a gel-like plug were evident at 10 days after feeding (Figure 1), indicating successful parasite development.
Next, to determine sandfly capacity to transmit the US strain of L. infantum to a susceptible vertebrate host, we allowed L. infantum–naive and L. infantum–infected sandflies to feed on 7 L. infantum–naive hamsters for 13 days. For confirmation of L. infantum infection, we dissected the alimentary tract of sandflies that fed on the hamsters (Table 2). A total of 30 sandflies were used for feeding on hamsters; 11 flies fed and were subjected to Leishmania detection by PCR, which confirmed Leishmania positivity for 5 sandflies. Hamster blood samples were collected 2 weeks after infection and monthly for 5 months. L. infantum DNA was detected in hamster blood by quantitative PCR (qPCR) and was present in hamster nos. 1 (at 2 mo), 2 (at 3 mo), 5 (at 4 mo), and 6 (at 4 mo) with cycle thresholds of 43.88, 28.27, 34.38, and 45 respectively. Cycle thresholds <45 were considered positive for L. infantum (5).
On hamster no. 5, a cutaneous lesion consistent with Leishmania infection persisted for 1 month. Tissue from this lesion was harvested to assay for Leishmania infection. Increased numbers of macrophages and granulocytes were present in the dermal layer. Bacteria found in the tissue probably represented secondary infection, a common sequela of canine VL. Cellular infiltrate was observed, indicative of inflammation and infection (Technical Appendix). No L. infantum parasites were observed on slides stained with hematoxylin and eosin, periodic acid–Schiff, or Giemsa, and lesion tissue was negative for L. infantum by qPCR (data not shown).
Figure 2

Figure 2. Visceralization of Leishmania infantum from US foxhounds, transmitted by sandflies into hamsters. Leishmania spp.–specific quantitative PCR was performed, and parasite load was calculated from a standard curve. Horizontal bars indicate mean...
qPCR was performed to quantify parasite load within common Leishmania-infected organs from all hamsters. The mean quantities of Leishmania DNA amplified from spleen, bone marrow, and lymph node from hamsters on which Leishmania-infected sandflies had fed were 12-, 22-, and 11-fold greater than that from hamsters on which Leishmania-naive sandflies had fed (Figure 2). According to extrapolation from a PCR standard curve similar to one previously used (5), the highest parasite load was in bone marrow, which contained an average of 1,238 (±282) parasites/mg tissue.
In the United States, parasites from VL-symptomatic, naturally infected hunting hounds remain highly infectious to Leishmania-competent Lu. longipalpis sandflies. Parasites that had fed on L. infantum–infected hounds were able to develop fully within sandflies and to be subsequently transmitted to and disseminated within hamsters.
The capacity of Lu. shannoni and Lu. longipalpis sandflies to acquire L. infantum from naturally infected dogs has been compared in leishmaniasis-endemic Brazil (13). Although lower infection rates were observed in Lu. shannoni (9%) than in Lu. longipalpis (36%) sandflies, the intensity of infection (200–500 promastigotes/fly) was higher in Lu. shannoni sandflies. This finding demonstrates a potentially lower threshold for acquiring infection from Lu. shannoni sandflies because the infectious dose per sandfly was greater. Lu. shannoni sandflies are commonly found within the United States and also in areas where L. infantum–infected hounds were reported (4). These data demonstrate the risk for vectorborne transmission of zoonotic VL from these dogs in the United States.
Despite our use of an L. infantum strain that is primarily, if not solely, transmitted via vertical transmission between dogs in the United States, we were able to measure substantial parasite loads in sandflies that fed on these dogs and in the bone marrow, spleen, and peripheral lymph nodes of hamsters on which infected sandflies had fed (Figures 1, 2). Parasite DNA was not amplified in the liver, possibly because of lower parasite loads in the liver during later infection, as demonstrated in experimental VL infections of mice (14). Therefore, the US strain of L. infantum that is circulating in North American hunting hounds has not lost virulence factors that facilitate adherence to sandfly gut and facilitate transmission, and subsequent dissemination, in a secondary host.
This study focused on the possibility that domestic hounds serve as reservoir hosts for L. infantum within the United States; however, other potential L. infantum reservoirs include coyotes, foxes, and opossums. When leishmaniasis was found to be reemerging among hounds in the United States in 2000, a total of 291 wild canids were trapped and tested (15). No serologic evidence of infection was found, but these studies were limited to the southeastern United States; further study is needed to rule out the possibility that enzootic cycles of transmission do not exist within wild canids. The range covered by Lu. shannoni sandflies overlaps that of reservoir species including coyotes, foxes, and hunting hounds. Occurrence of Leishmania vectors in areas of naturally infected hounds indicates a coalescence of components for establishment of a sylvatic and/or domestic cycle of L. infantum. Diagnostic testing and preventive measures should be considered for dog breeds known to harbor L. infantum.
In the United States, L. infantum is circulating among dogs. Despite the fact that vertical transmission maintains VL within the hound population (5), L. infantum was able to fully develop in sandflies and be further transmitted to a susceptible vertebrate host. Symptomatic hounds were highly infectious to sandflies. L. infantum strain MCAN/US/2001/FOXYMO1, similar to the common European zymodeme MON-1, circulating dog-to-dog in North America maintained all necessary requirements for complete development within sandflies. Overlap of sandfly infections (e.g., Lu. shannoni, and L. infantumi) in hounds may put companion dogs and humans at risk and could pose an emerging risk for L. infantum–triggered clinical disease in at-risk populations in North America.
Dr. Schaut is a postdoctoral fellow who works in the laboratory of C.A.P. at the University of Iowa. His research focuses on immunologic and other alterations that occur during chronic VL.
Acknowledgments
We thank members of the laboratory of C.A.P. for helpful suggestions; Douglas Jones for troubleshooting assistance; Chembio Diagnostic Systems, Inc. (Medford, NY, USA) for donation of Leishmania dual-path platform tests; collaborating foxhunting clubs for use of their priceless hounds; and Iowa State University Laboratory Animal Resources for valuable assistance.
This work was funded by grant AI088051 from the National Institutes of Allergy and Infectious Diseases, National Institutes of Health, to M.R.O. and C.A.P. K.J.E. was funded by a fellowship from the Morris Animal Foundation and Pfizer.
References
- Alvar J, Velez ID, Bern C, Herrero M, Desjeux P, Cano J, Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE. 2012;7:e35671. DOIPubMedGoogle Scholar
- Ashford DA, David JR, Freire M, David R, Sherlock I, Eulalio MC, Studies on control of visceral leishmaniasis: impact of dog control on canine and human visceral leishmaniasis in Jacobina, Bahia, Brazil. Am J Trop Med Hyg. 1998;59:53–7 .PubMedGoogle Scholar
- Anderson DC, Buckner RG, Glenn BL, MacVean DW. Endemic canine leishmaniasis. Vet Pathol. 1980;17:94–6 .PubMedGoogle Scholar
- Petersen CA, Barr SC. Canine leishmaniasis in North America: emerging or newly recognized? Vet Clin North Am Small Anim Pract. 2009;39:1065–74. DOIPubMedGoogle Scholar
- Boggiatto PM, Gibson-Corley KN, Metz K, Gallup JM, Hostetter JM, Mullin K, Transplacental transmission of Leishmania infantum as a means for continued disease incidence in North America. PLoS Negl Trop Dis. 2011;5:e1019. DOIPubMedGoogle Scholar
- Gibson-Corley KN, Hostetter JM, Hostetter SJ, Mullin K, Ramer-Tait AE, Boggiatto PM, Disseminated Leishmania infantum infection in two sibling foxhounds due to possible vertical transmission. Can Vet J. 2008;49:1005–8 .PubMedGoogle Scholar
- Chattopadhayay R, Kaur S, Ganguly NK, Mahajan RC. Antigenic differences between axenic amastigotes & promastigotes of Leishmania donovani. Indian J Med Res. 1996;104:349–54 .PubMedGoogle Scholar
- Petersen CA. Leishmaniasis, an emerging disease found in companion animals in the United States. Top Companion Anim Med. 2009;24:182–8. DOIPubMedGoogle Scholar
- Clarke CF, Bradley KK, Wright JH, Glowicz J. Case report: emergence of autochthonous cutaneous leishmaniasis in northeastern Texas and southeastern Oklahoma. Am J Trop Med Hyg. 2013;88:157–61. DOIPubMedGoogle Scholar
- Douvoyiannis M, Khromachou T, Byers N, Hargreaves J, Murray HW. Cutaneous leishmaniasis in North Dakota. Clin Infect Dis. 2014;59:e73–5. DOIPubMedGoogle Scholar
- Coutinho-Abreu IV, Sonoda IV, Fonseca JA, Melo MA, Balbino VQ, Ramalho-Ortigao M. Lutzomyia longipalpis s.l. in Brazil and the impact of the Sao Francisco River in the speciation of this sand fly vector. Parasit Vectors. 2008;1:16.
- Weng JL, Young SL, Gordon DM, Claborn D, Petersen C, Ramalho-Ortigao M. First report of phlebotomine sandflies (Diptera: Psychodidae) in Kansas and Missouri, and a PCR method to distinguish Lutzomyia shannoni from Lutzomyia vexator. J Med Entomol. 2012;49:1460–5. DOIPubMedGoogle Scholar
- Travi BL, Ferro C, Cadena H, Montoya-Lerma J, Adler GH. Canine visceral leishmaniasis: dog infectivity to sandflies from non-endemic areas. Res Vet Sci. 2002;72:83–6. DOIPubMedGoogle Scholar
- Ahmed S, Colmenares M, Soong L, Goldsmith-Pestana K, Munstermann L, Molina R, Intradermal infection model for pathogenesis and vaccine studies of murine visceral leishmaniasis. Infect Immun. 2003;71:401–10. DOIPubMedGoogle Scholar
- Duprey ZH, Steurer FJ, Rooney JA, Kirchhoff LV, Jackson JE, Rowton ED, Canine visceral leishmaniasis, United States and Canada, 2000–2003. Emerg Infect Dis. 2006;12:440–6. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 21, Number 12—December 2015
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Christine A. Petersen, University of Iowa Department of Epidemiology, S429 CPHB, 145 N Riverside Dr, Iowa City, IA 52241, USA
Top