Volume 21, Number 3—March 2015
Dispatch
Comparison of Porcine Epidemic Diarrhea Viruses from Germany and the United States, 2014
Cite This Article
Citation for Media
Abstract
Since 2013, highly virulent porcine epidemic diarrhea virus has caused considerable economic losses in the United States. To determine the relation of US strains to those recently causing disease in Germany, we compared genomes and found that the strain from Germany is closely related to variants in the United States.
Porcine epidemic diarrhea (PED) is an acute and highly contagious enteric disease of swine that results in severe enteritis, diarrhea, vomiting, and dehydration (1). Porcine epidemic diarrhea virus (PEDV), the causative agent, is an enveloped, positive single-stranded RNA virus that belongs to the family Coronaviridae, genus Alphacoronavirus (2).
The disease was first recognized in Europe in 1971 and has thereafter caused high economic losses, particularly in Asia. In May 2013, a highly virulent PEDV variant emerged in the United States; explosive epidemics on swine farms affected pigs of all ages, resulting in a mortality rate of up to 95% among suckling pigs (3). Since then, outbreaks have occurred in 30 US states (4), causing very high economic losses, and the disease threatens to spread. The involved viruses cluster together with isolates from China from 2011 and 2012 (5). Apart from these highly virulent strains, a PEDV variant from the United States (strain OH851) that affected sows instead of younger animals and caused milder disease was recently described (6).
The effect of PED in the United States has unsettled pig farmers and veterinarians worldwide; studies have been recently initiated to elucidate the situation in Europe. Despite the history of PED outbreaks in Europe, little is known about currently circulating virus strains and their effect; information about the phylogeny of recent strains and their relation to the outbreak strain in the United States is lacking.
We report a case of PED that occurred on a swine-fattening farm in Germany in May 2014. The causative virus was fully characterized by using conventional methods and next-generation sequencing. We analyzed the resulting full-length genomes and compared them with those of the strains circulating in the United States and Asia to elucidate possible relationships.
Figure 1
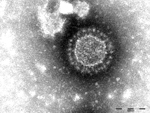
Figure 1. Porcine epidemic diarrhea virus particles seen by negative-stain electron microscopy of fecal samples. Negative staining with 1% phosphotungstic acid. Scale bar indicates 100 nm.
In May 2014, a pig fattening farm in southern Germany (Federal State of Baden-Wuerttemberg) that continuously houses ≈1,400 fattening pigs reported watery diarrhea in pigs in all age groups (feeders to slaughter animals). The first cases occurred after new feeder pigs from a large piglet producer were brought onto the farm. The incoming animals showed diarrhea 2 days after arrival. Within 1 day thereafter, the disease had spread to pigs in all other age groups. Clinical signs were present for at least 1 week; ≈20 pigs died. Fecal samples from diseased pigs were submitted to the regional laboratory for diagnosis, and coronaviruses were detected by electron microscopy (Figure 1). Additionally, 3 pigs with catarrhal enteritis were euthanized; postmortem examination at the regional laboratory confirmed coronavirus infection in all 3 animals.
Subsequently, PED was diagnosed in a private laboratory (IVD GmbH, Hannover, Germany) by use of a published multiplex reverse transcription quantitative PCR (7). Selected positive samples were submitted to the Friedrich-Loeffler-Institut, Isle of Riems, Germany, for confirmation and further virus characterization. At this institution, 2 fecal samples with high genome load, as determined by 2 independent, published (8,9) reverse transcription quantitative PCRs selective for PEDV nucleocapsid (N) and spike (S) genes, were chosen for routine virologic testing and next-generation sequencing. Sequencing and data analyses were performed as previously described (10). The phylogenetic tree, based on full-length genomes, was constructed by using PhyML (11) in the Geneious software suite (http://www.geneious.com/) with a generalized time reversible substitution model, and the tree was supported by 1,000 bootstrapping replicates.
Virus isolates were obtained after inoculation of cells of different permanent cell lines (pig kidney [PK]-15 and Vero) with clinical material from the pigs. Sequencing of nucleic acids isolated directly from diagnostic samples PEDV/GER/L00719/2014 and PEDV/GER/L00721/2014 resulted in 2 viral genomes (Table) encompassing all typical PEDV coding sequences. Each sequence encodes a large replicase polyprotein, a spike (S) protein, an alphacoronavirus-specific accessory membrane protein, an envelope protein, a membrane protein, and a nucleocapsid (N) protein.
Comparative analyses of the full genomes showed that the strains share a very high (99.5%) identity with the new variant OH851 (GenBank accession no. KJ399978) recently reported from the United States (6). A more comprehensive comparison of 21 full-length PEDV genomes from different years and locations revealed lower similarities (≈98.7%) with currently circulating highly virulent strains from the United States and from China (Technical Appendix Figure, panel A). In contrast, the new isolates from Germany, PEDV/GER/L00719/2014 and PEDV/GER/L00721/2014, are less similar (97.1%) to the isolate from Europe, CV777, which dates back to the 1970s (12).
The nucleotide alignment of the obtained PEDV genomes and the available references from the database revealed a region with high variability of the first 1,200 nt in the 5′ portion of the S protein–coding sequences (Technical Appendix Figure, panel B). The N terminal S1 domain of the coronavirus S protein is necessary for virus attachment by interaction with host cell receptors (13) and might therefore be highly mutable.
Although in-depth analysis of the deep-sequencing data for PEDV/GER/L00721/2014 revealed a genetically homogenous population, this analysis for PEDV/GER/L00719/2014 uncovered a mixed viral population with a total of 8 single-nucleotide variants. One nonsynonymous single-nucleotide variant (variant position G19042U, amino acid substitution S6348I) was detected in the polyprotein coding sequences. Of note, 7 single-nucleotide variants are located in the aforementioned variable region in the S protein coding sequences, 5 of which are nonsynonymous (Table), thereby confirming the high variability in the N terminal part of the S protein.
Figure 2

Figure 2. Phylogenetic analysis based on 21 full-length porcine epidemic diarrhea virus (PEDV) genomes. The new strains from Germany (PEDV/GER/L00719/2014 and PEDV/GER/L00721/2014, in boldface and italics) and the new 2014 PEDV variant from...
Because quite extensive differences (≈50 aa) were found between the recent N terminal S protein region of the isolates from Germany and the highly virulent PEDV strains from the United States and China, the isolates from Germany described in this article seem to not be directly linked to the highly virulent PEDV strains circulating in the United States (Figure 2). In contrast, the recent isolates from Germany and strain OH851 share not only high identity over the entire genome, including the highly variable 5′ region of the S protein coding sequences, but also their clinical phenotype observed under field conditions.
PEDV infection was confirmed in a pig herd in Germany in 2014. Comparative analyses of full-length sequences revealed that the isolates from these pigs in Germany show very high nucleotide similarity with strain OH851 found in the United States in 2014. However, differences exist that distinguish the strains from Germany from the highly virulent PEDV strains that caused the major losses in the United States. Given the fact that PEDV surveillance has been lacking in Germany, we cannot exclude the possibility that the strains described here have already been circulating in Europe for a longer time or were indeed recently introduced from the United States or Asia to Europe. Therefore, our report provides useful information about recent PEDV strains in Europe, but a comprehensive evaluation is still difficult because of a lack of data about additional strains. Future studies should therefore concentrate on analysis of additional PEDV from different years and locations.
Mr. Hanke and Mrs. Jenckel are doctoral students at the Institute of Diagnostic Virology, Friedrich-Loeffler-Institut, Isle of Riems, Germany. Their research focuses on the rational use of next-generation sequencing in metagenomic analyses and molecular epidemiology.
Acknowledgments
We thank all technicians involved in the study.
The study was financially supported by the German Ministry of Nutrition and Agriculture and by the European Research Area project EpiSeq (ERANET, htttp://www.episeq.eu/).
References
- Song D, Park B. Porcine epidemic diarrhoea virus: a comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes. 2012;44:167–75 . DOIPubMedGoogle Scholar
- International Committee on Taxonomy of Viruses. Virus taxonomy: classification and nomenclature of viruses; ninth report of the International Committee on Taxonomy of Viruses. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ, editors. San Diego: Elsevier Academic Press; 2012.
- Stevenson GW, Hoang H, Schwartz KJ, Burrough ER, Sun D, Madson D, Emergence of porcine epidemic diarrhea virus in the United States: clinical signs, lesions, and viral genomic sequences. J Vet Diagn Invest. 2013;25:649–54. DOIPubMedGoogle Scholar
- US Department of Agriculture. Swine enteric coronavirus disease testing summary report. [cited 2014 Nov 21]. http://www.aphis.usda.gov/animal_health/animal_dis_spec/swine/downloads/SECD_pre-fed_order_nahln_test_sum_rpt.pdf
- Chen Q, Li G, Stasko J, Thomas JT, Stensland WR, Pillatzki AE, Isolation and characterization of porcine epidemic diarrhea viruses associated with the 2013 disease outbreak among swine in the United States. J Clin Microbiol. 2014;52:234–43. DOIPubMedGoogle Scholar
- Wang L, Byrum B, Zhang Y. New variant of porcine epidemic diarrhea virus, United States, 2014. Emerg Infect Dis. 2014;20:917–9. DOIPubMedGoogle Scholar
- Kim SH, Kim IJ, Pyo HM, Tark DS, Song JY, Hyun BH. Multiplex real-time RT-PCR for the simultaneous detection and quantification of transmissible gastroenteritis virus and porcine epidemic diarrhea virus. J Virol Methods. 2007;146:172–7. DOIPubMedGoogle Scholar
- University of Minnestota. New rapid semi-quantitative RT-PCR assay developed to detect porcine epidemic diarrhea virus [cited 2014 Nov 21]. http://www.cahfs.umn.edu/prod/groups/cvm/@pub/@cvm/@cahfs/documents/content/cvm_content_446868.pdf
- Alonso C, Goede DP, Morrison RB, Davies PR, Rovira A, Marthaler DG, Evidence of infectivity of airborne porcine epidemic diarrhea virus and detection of airborne viral RNA at long distances from infected herds. Vet Res. 2014;45:73. DOIPubMedGoogle Scholar
- Juozapaitis M, Moreira É, Mena I, Giese S, Riegger D, Pohlmann A, An infectious bat-derived chimeric virus harbouring the entry machinery of an influenza A virus. Nat Commun. 2014;5.4448. PMID:25055345 DOIGoogle Scholar
- Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol. 2010;59:307–21. DOIPubMedGoogle Scholar
- Debouck P, Pensaert M. Experimental infection of pigs with a new porcine enteric coronavirus, CV 777. Am J Vet Res. 1980;41:219–23 .PubMedGoogle Scholar
- Bosch BJ, van der Zee R, de Haan CA, Rottier PJ. The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. J Virol. 2003;77:8801–11. DOIPubMedGoogle Scholar
Figures
Table
Cite This Article1These authors contributed equally to this article.
Table of Contents – Volume 21, Number 3—March 2015
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Sandra Blome, Friedrich-Loeffler-Institut, Institute of Diagnostic Virology, Südufer 10, 17493 Greifswald–Insel Riems, Germany
Top