Volume 21, Number 4—April 2015
Dispatch
Multidrug-Resistant Salmonella enterica Serotype Typhi, Gulf of Guinea Region, Africa
Cite This Article
Citation for Media
Abstract
We identified 3 lineages among multidrug-resistant (MDR) Salmonella enterica serotype Typhi isolates in the Gulf of Guinea region in Africa during the 2000s. However, the MDR H58 haplotype, which predominates in southern Asia and Kenya, was not identified. MDR quinolone-susceptible isolates contained a 190-kb incHI1 pST2 plasmid or a 50-kb incN pST3 plasmid.
Typhoid fever, which is caused by Salmonella enterica serotype Typhi, is endemic to the developing world; there were an estimated 26.7 million cases in 2010 (1). The incidence of typhoid fever in sub-Saharan Africa was an estimated 725 cases/100,000 persons in 2010, despite a lack of incidence studies conducted in West and central Africa (1). Antimicrobial susceptibility data are also scarce for this part of Africa. This issue is problematic because treatment with appropriate antimicrobial drugs is essential for recovery in the context of the global emergence of multidrug resistance.
In the Indian subcontinent and Southeast Asia, the multidrug-resistant (MDR) Salmonella Typhi H58 clone, which was named after its haplotype (a combination of defined chromosomal single-nucleotide polymorphisms [SNPs]) (2,3), has spread rapidly and become endemic and predominant. During the 1990s, this clone acquired a large conjugative incHI1 pST6 plasmid encoding resistance to ampicillin, chloramphenicol, and co-trimoxazole (4,5); also in the 1990s, this MDR clone became resistant to quinolones and showed decreased susceptibility to ciprofloxacin because of point mutations in the chromosomal gyrA gene (2). The H58 clone has also spread to eastern Africa, where it has been the most prevalent haplotype (87%) in Kenya since the early 2000s (6).
During 1997–2011, high incidence of MDR Salmonella Typhi was reported in some countries near the Gulf of Guinea in Africa, including Nigeria (7), Ghana (8,9), Togo (10), and the Democratic Republic of the Congo (11). During 1999–2003, a British surveillance system reported a prevalence of 19% (49/421) for MDR Salmonella Typhi isolates among imported cases of typhoid fever acquired in Africa, particularly in Ghana (12). However, nothing is known about the genotypes of these isolates, including whether they belong to the spreading MDR H58 clone.
We report data for the occurrence, genotypes, and characterization of the resistance mechanisms of MDR Salmonella Typhi isolates. These isolates were obtained from the French National Reference Center for Salmonella (FNRC-Salm), Institut Pasteur (Paris, France), and Centre Pasteur du Cameroun (Yaoundé, Cameroon).
Almost all Salmonella Typhi strains isolated in France are referred to the FNRC-Salm. Most isolates were obtained from travelers or immigrants, most of whom were infected in Africa and Asia. In Cameroon, the Centre Pasteur du Cameroun collects Salmonella Typhi isolates from several hospitals in Yaoundé.
Antimicrobial susceptibility testing was performed according to the guidelines of the antibiogram committee of the French Society for Microbiology (http://www.sfm.asso.fr/nouv/general.php?pa = 2). Isolates were considered to be MDR if they were resistant to ≥2 of the following antimicrobial drugs: amoxicillin, co-trimoxazole (trimethoprim/sulfamethoxazole), chloramphenicol, or tetracyclines.
During 1996–2013, a total of 1,746 Salmonella Typhi isolates were collected through the French national surveillance system and subjected to antimicrobial susceptibility testing; 408 were acquired in sub-Saharan Africa (n = 237) and northern Africa (n = 171), and 55 (13.5%) of those acquired in Africa were MDR (Table). All but 1 of the MDR isolates were acquired in sub-Saharan Africa (Table). The proportion of MDR isolates increased from 0% during 1996–1999 to 22.3% (n = 23) during 2010–2013. Only 4 isolates from Africa were resistant to nalidixic acid, including 1 isolate resistant to ciprofloxacin. Because these isolates acquired after 2010 were not MDR isolates, they were not studied further.
In Yaoundé, the proportion of MDR isolates was high (45.5 %, 29/61) in the first survey during 2000–2004. However, this proportion increased to 75.5% (37/49) during 2010–2013.
We studied 61 isolates (Technical Appendix). Of these, 46 were MDR: 29 acquired in Africa and detected at FNRC-Salm before 2010; 2 acquired in France during 2009 at an African festive meal (13); 12 randomly selected acquired in Yaoundé during 2002–2007; 2 acquired in the Central African Republic, and 1 acquired in Morocco (2). The remaining 15 comparison strains (MDR or drug susceptible) that belonged to various haplotypes and were acquired in Africa and Asia during 1958–2009.
Mechanisms of antimicrobial drug resistance were determined as described (14). Genetic diversity and phylogenetic relationships were studied by using standardized XbaI–pulsed-field gel electrophoresis (PFGE) (http://www.cdc.gov/pulsenet/pathogens/index.html), haplotyping (5), and clustered regularly interspaced short palindromic repeats (CRISPR) typing (15). Haplotyping was based on identification of SNPs at 1,487 defined chromosomal loci, and CRISPR typing was based on detection of 32-bp sequences (spacers) within 1 or both CRISPR regions.
Figure 1

Figure 1. Characteristics of 50 Salmonella enterica serotype Typhi isolates. The dendrogram was generated by using BioNumerics version 6.6 software (Applied Maths, Sint-Martens-Latem, Belgium) and shows results of cluster analysis on the basis...
Figure 2
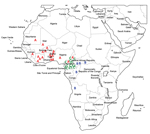
Figure 2. Distribution of multidrug-resistant Salmonella enterica serotype Typhi isolates by genetic lineage (A, B, or C), Gulf of Guinea region, Africa. Location within the country of infection/isolation was assigned at random. UAE,...
With the exception of the isolate from Morocco (ISP-03-07467) (2), none of the isolates had the H58 haplotype or contained the associated incHI1 pST6 MDR plasmid. We found 3 other lineages with different geographic distributions and MDR plasmids (Figures 1, 2).
Lineage A consisted mostly of haplotype H56 isolates and more rarely H42 (which differs from H56 by 2 SNPs) and was found only in the western part of the Gulf of Guinea region. Lineage B consisted of haplotype H55 isolates and was found in the eastern and southern parts of the Gulf of Guinea region. Lineage C consisted of haplotype H77 isolates and was found only in Cameroon. All 3 lineages had distinctive CRISPR1 spacer contents. XbaI-PFGE, which used a similarity value of ≥90% as a cutoff, correctly grouped (i.e., concordant with haplotyping and CRISPR results) all but 2 of the MDR isolates from Africa (Figure 1; Technical Appendix).
The 3 lineages contained a large (≈190 kb) conjugative MDR incHI1 pST2 plasmid that differed among lineages. Resistance to trimethoprim was encoded by different class 1 integron gene cassettes: dfrA15, dfrA7, and dfrA1 for incHI1 plasmids of lineages A, B, and C, respectively. All incHI1 plasmids from lineage A encoded resistance to chloramphenicol, and none of those from lineage C encoded such resistance. A second smaller (50-kb) MDR plasmid belonging to the incN incompatibility group (pST3 by plasmid multilocus sequence typing), was present mostly in lineage C isolates, but was also found in 1 lineage A isolate (02-1739) (Technical Appendix).
Analysis of older isolates and previously published data (2) showed that susceptible Salmonella Typhi strains of haplotypes H42, H56, and H77 had already been identified in Senegal in 1962, Tunisia in 1969, and Cameroon in 1958, respectively. This finding suggests that the MDR isolates from lineages A and C are derived from local Salmonella Typhi populations in Africa, rather than being recently imported from other regions to which this bacterium is endemic. Haplotype H55 was previously restricted largely to the Indian subcontinent and eastern Africa (2); it was detected in association with an incHI1 pST2 plasmid in India during the mid-1970s (5). Therefore, lineage B might have been was imported into central Africa from eastern Africa/southern Asia.
A previous study also reported isolation of an MDR clone in the Democratic Republic of the Congo in 2004 that was resistant to quinolones, showed decreased susceptibility to ciprofloxacin, and belonged to the Asian H58 lineage (2). Because only a limited number of isolates from central Africa were tested in our study, studies of a larger collection of isolates might provide more information about bacterial genotypes/MDR plasmids circulating in central Africa.
Despite intrinsic limitations of a laboratory surveillance system for typhoid fever that is used mostly for travelers and immigrants and has the bias of preferential links caused by colonial history and choices of tourist destinations, we documented emergence of 3 MDR Salmonella Typhi lineages in the Gulf of Guinea area. Two lineages found in Guinea and Cameroon were local lineages that acquired MDR conjugative plasmids, either a large incHI1 pST2 plasmid or a smaller incN pST3 plasmid. The H58 lineage, which is currently predominant in Asia and eastern Africa, was not detected among MDR isolates from West and central Africa.
At the time of this study, Ms. Baltazar was a predoctoral student at the FNRC-Salm, Paris, France. She is currently a doctoral candidate at the University of Limoges, Limoges, France. Her primary research interest is mechanisms of antimicrobial drug resistance.
Acknowledgments
We thank all the corresponding laboratories of the FNRC-Salm network for participating in this study.
This study was supported by the Institut Pasteur, the Réseau International des Instituts Pasteur, the Institut de Veille Sanitaire, and the French Government Investissement d'Avenir Program (Integrative Biology of Emerging Infectious Diseases, Laboratory of Excellence, grant ANR-10-LABX-62-IBEID).
References
- Buckle GC, Walker CL, Black RE. Typhoid fever and paratyphoid fever: systematic review to estimate global morbidity and mortality for 2010. J Glob Health. 2012;2:010401. DOIPubMedGoogle Scholar
- Roumagnac P, Weill FX, Dolecek C, Baker S, Brisse S, Chinh NT, Evolutionary history of Salmonella Typhi. Science. 2006;314:1301–4. DOIPubMedGoogle Scholar
- Holt KE, Parkhill J, Mazzoni CJ, Roumagnac P, Weill FX, Goodhead I, High-throughput sequencing provides insights into genome variation and evolution in Salmonella Typhi. Nat Genet. 2008;40:987–93. DOIPubMedGoogle Scholar
- Phan MD, Kidgell C, Nair S, Holt KE, Turner AK, Hinds J, Variation in Salmonella enterica serovar Typhi IncHI1 plasmids during the global spread of resistant typhoid fever. Antimicrob Agents Chemother. 2009;53:716–27. DOIPubMedGoogle Scholar
- Holt KE, Phan MD, Baker S, Duy PT, Nga TV, Nair S, Emergence of a globally dominant IncHI1 plasmid type associated with multiple drug resistant typhoid. PLoS Negl Trop Dis. 2011;5:e1245. DOIPubMedGoogle Scholar
- Kariuki S, Revathi G, Kiiru J, Mengo DM, Mwituria J, Muyodi J, Typhoid in Kenya is associated with a dominant multidrug-resistant Salmonella enterica serovar Typhi haplotype that is also widespread in Southeast Asia. J Clin Microbiol. 2010;48:2171–6. DOIPubMedGoogle Scholar
- Akinyemi KO, Coker AO. Trends of antibiotic resistance in Salmonella enterica serovar typhi isolated from hospitalized patients from 1997 to 2004 in Lagos, Nigeria. Indian J Med Microbiol. 2007;25:436–7. DOIPubMedGoogle Scholar
- Mills-Robertson F, Addy ME, Mensah P, Crupper SS. Molecular characterization of antibiotic resistance in clinical Salmonella typhi isolated in Ghana. FEMS Microbiol Lett. 2002;215:249–53. DOIPubMedGoogle Scholar
- Gross U, Amuzu SK, de Ciman R, Kassimova I, Gross L, Rabsch W, Bacteremia and antimicrobial drug resistance over time, Ghana. Emerg Infect Dis. 2011;17:1879–82. DOIPubMedGoogle Scholar
- Dagnra AY, Akolly K, Gbadoe A, Aho K, David M. Emergence of multidrug resistant Salmonella strains in Lome (Togo) [in French]. Med Mal Infect. 2007;37:266–9. DOIPubMedGoogle Scholar
- Lunguya O, Lejon V, Phoba MF, Bertrand S, Vanhoof R, Verhaegen J, . Salmonella Typhi in the Democratic Republic of the Congo: fluoroquinolone decreased susceptibility on the rise. EPLoS Negl Trop Dis. 2012; 6:e1921. Doe: . Pub 2012 Nov 5.DOIGoogle Scholar
- Cooke FJ, Day M, Wain J, Ward LR, Threlfall EJ. Cases of typhoid fever imported to England, Scotland and Wales (2000–2003). Trans R Soc Trop Med Hyg. 2007;101:398–404. DOIPubMedGoogle Scholar
- Loury P, Tillaut H, Faisant M, Paillereau N, Marquis M, Mari C, Cluster of typhoid fever cases in Ille-et-Vilaine (France), April 2009 [in French]. Bull Epidémiol Hebd. 2010;44:446–8.
- Le Hello S, Harrois D, Bouchrif B, Sontag L, Elhani D, Guibert V, Highly drug-resistant Salmonella enterica serotype Kentucky ST198-X1: a microbiological study. Lancet Infect Dis. 2013;13:672–9. DOIPubMedGoogle Scholar
- Fabre L, Le Hello S, Roux C, Issenhuth-Jeanjean S, Weill FX. CRISPR is an optimal target for the design of specific PCR assays for Salmonella enterica serotypes Typhi and Paratyphi A. PLoS Negl Trop Dis. 2014;8:e2671 . DOIPubMedGoogle Scholar
Figures
Table
Cite This Article1These authors contributed equally to this article.
2Current affiliation: University of Limoges, Limoges, France.
3Current affiliation: Rockefeller University, New York, New York, USA.
4Current affiliation: Institute Pasteur de Dakar, Dakar, Senegal.
Table of Contents – Volume 21, Number 4—April 2015
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
François-Xavier Weill, Centre National de Référence des Escherichia coli, Shigella et Salmonella, Unité des Bactéries Pathogènes Entériques, Institut Pasteur, 28 Rue du Docteur Roux, 75724 Paris CEDEX 15, France
Top