Volume 22, Number 12—December 2016
Research
Cutaneous Granulomas in Dolphins Caused by Novel Uncultivated Paracoccidioides brasiliensis
Cite This Article
Citation for Media
Abstract
Cutaneous granulomas in dolphins were believed to be caused by Lacazia loboi, which also causes a similar disease in humans. This hypothesis was recently challenged by reports that fungal DNA sequences from dolphins grouped this pathogen with Paracoccidioides brasiliensis. We conducted phylogenetic analysis of fungi from 6 bottlenose dolphins (Tursiops truncatus) with cutaneous granulomas and chains of yeast cells in infected tissues. Kex gene sequences of P. brasiliensis from dolphins showed 100% homology with sequences from cultivated P. brasiliensis, 73% with those of L. loboi, and 93% with those of P. lutzii. Parsimony analysis placed DNA sequences from dolphins within a cluster with human P. brasiliensis strains. This cluster was the sister taxon to P. lutzii and L. loboi. Our molecular data support previous findings and suggest that a novel uncultivated strain of P. brasiliensis restricted to cutaneous lesions in dolphins is probably the cause of lacaziosis/lobomycosis, herein referred to as paracoccidioidomycosis ceti.
The clinical and phenotypic features of the uncultivated agent of lacaziosis/lobomycosis in dolphins suggested that this pathogen was the same organism as Lacazia loboi, which causes skin keloidal-like lesions in humans (1–6). Although several studies indicated that L. loboi from human resists culture (4–6), only 1 well-documented study shows the uncultivated nature of the pathogen causing cutaneous granulomas in dolphins (7). Thus, the true ecology, epidemiology, and taxonomy of these 2 uncultivated pathogens of humans and dolphins have been controversial (4,7).
Because of their phenotypic resemblance and serologic cross-reactivity with Paracoccidioides brasiliensis, at one time these pathogens were believed to be P. loboi (4,8). This taxonomic controversy was partially resolved in 1999 when Taborda et al. (9) proposed the binomial L. loboi and concluded that previous terms used to name the etiologic agent of skin keloidal-like lesions in humans and dolphins were invalid. Molecular analysis of internal transcriber spacer (ITS) and chitin synthase 4 (CHS4) genes validated their original proposal (10). Further phylogenetic analysis of several genomic DNA sequences showed that L. loboi was closely related to Paracoccidioides spp. (11). However, other molecular data showed that L. loboi from humans was located in its own genus because of strong bootstrap support (12).
The notion that human L. loboi was the same organism as those in the skin of dolphins with lacaziosis/lobomycosis was first challenged by Rotstein et al. (13), who used molecular analysis. These investigators found that the 28S rDNA amplicon of L. loboi in extracted genomic DNA from an infected bottlenose dolphin (Tursiops truncatus) in North Carolina, USA, coastal areas had 97% identity with P. brasiliensis DNA sequences available in GenBank. However, their DNA sequences are not available. More recently, 3 groups in Japan (14,15) and Spain (16), who also used molecular methods, reported similar observations for several dolphin species including, T. truncatus and Lagenorhynchus obliquidens, which had skin granulomas and yeast-like cells in infected tissues. These studies showed that glycoprotein 43 (gp43)–like and ITS partial DNA sequences isolated from infected dolphins placed the etiologic agent of skin granulomas among human P. brasiliensis strains.
We amplified by using PCR the partial coding DNA sequences of the Kex gene in genomic DNA isolated from 6 bottlenose dolphins with cutaneous granulomas. These dolphins were captured in the Indian River Lagoon, Florida, USA, a 156-mile estuary along the eastern coast of the United States. Phylogenetic analysis showed that Kex PCR amplicons, which contained partial DNA sequences of the Kex protein, clustered among cultivated P. brasiliensis strains from humans with systemic paracoccidioidomycosis. Our data suggest that a novel uncultivated P. brasiliensis type, different from L. loboi from humans, is the probable etiologic agent of cutaneous granulomas in dolphins.
Biopsy Specimens from Bottlenose Dolphins
Four formalin-fixed tissues were received from the Harbor Branch Oceanographic Institute (Fort Pierce, FL, USA). Samples were collected in June 2003 from bottlenose dolphins captured in the Indian River Lagoon with cutaneous granulomas displaying chains of yeast cells in the infected tissues (FB 921, FB938, FB946, and FB952). Two additional skin biopsy specimens were obtained from SeaWorld of Texas (San Antonio, TX, USA); 1 specimen (SW070458) was collected during rescue and rehabilitation efforts, and a second 1 specimen (B92-932) was obtained from an animal that came from the Indian River Lagoon and was then kept at SeaWorld of Texas (Table).
Isolation of DNA from Paraffin-Embedded Tissues
Using a sterile microtome, we obtained 10-mm–thick sections from paraffin-embedded tissues. Parts of sections were examined by using histopathologic analysis after staining with Gomori methenamine silver to verify the presence and quantity of yeast-like cells in selected specimens.
Isolation of DNA was performed by using the BioChain FFPET protocol (BioChain Institute, Inc., Newark, CA, USA). In brief, at least three 10-mm–thick sections were placed in a 1.5-mL microcentrifuge tube, and 500 μL of Dewaxil reagent was added. The sample was incubated at 90°C for 1 h, followed by addition of 180 μL of lysis buffer and a brief centrifugation. Two phases were formed; 20 μL of proteinase K was added to the lower phase, and the mixture was incubated at 56°C for 1.5 h. After incubation, the sample was centrifuged for 1 min, and the lower phase was transferred into a new tube. RNase A (2.0 μL, 100 mg/mL) was added, followed by addition of 100 μL of binding buffer and 100 μL of 100% ethanol. The entire mixture was then transferred into a separation column (BioChain Institute, Inc.) and centrifuged at 6,000 × g for 1 min. The column was washed twice with the provided buffers. DNA was extracted by adding 50 μL of elution buffer and centrifuging for 1 min at maximum speed. Samples were used immediately or stored at −80°C.
Amplification and Sequencing of Partial Kex Gene Sequences
Because genomic DNA extraction from formalin-fixed tissues usually degrades genome DNA into small pieces, we designed primers targeting fragments <300 bp. To properly verify previous findings, we selected a conserved region of the Kex partial DNA sequence to target a DNA epitope other than gp43 and ITS sequences used by other investigators (14–16). We used the protocol of Vilela et al. (12) to search for homologous DNA sequences of Kex protein in GenBank, aligned sequences by using ClustalW, version 1.81 (17), and inspected them visually.
Conserved regions were selected to construct the set of primers Kex-1F 5′-TGCTTYGGTTTGGGGTTG-3′ and Kex-2R 5′-CACTGGARCCGTCAGCTA-3′. The set of primers were designed to amplify a 151-bp region of the Kex DNA sequence according to the PCR protocol of Vilela et al. (12). Amplicons were ligated into the pCR 2.1-TOPO vector (Invitrogen, Carlsbad, CA. USA), purified, and sequenced by using BigDye Terminator Chemistry in an ABI Prim 310 Genetic Analyzer (Applied Biosystems, Foster City, CA. USA).
To further corroborate our results, we used gp43 DNA sequences reported by Minakawa et al. (GenBank accession no. AB811031) (14) and Ueda et al. (GenBank accession no. LC067206) (15) and ITS DNA sequences reported by Esperón et al. (GenBank accession no. HQ413323) (16) for phylogenetic analysis of several homologous DNA sequences of P. brasiliensis, P. lutzii, and L. loboi in GenBank. We also analyzed 2 unpublished CHS4 gene sequences (GenBank accession nos. KX267767 [A3] and KX267768 [90A]; A. Schaefer, P, McCarthy, unpub. data) isolated in 2008 from 2 dolphins with lacaziosis/lobomycosis in the Indian River Lagoon.
Phylogenetic Analyses
Homologous DNA sequences of partial CHS4, gp43, Kex, and ITS sequences of P. brasiliensis, P. lutzii, L. loboi, Ajellomyces capsulatus, and A. dermatitidis were aligned by using default settings in ClustalW, version 1.81 (17) inspected visually, and exported for analysis by using maximum-parsimony and neighbor-joining in MEGA6 (http://www.megasoftware.net) (18). Aligned sequences were exported for parsimony analysis by using a heuristic search with tree bisection reconnection branch swapping (MEGA6) and distant analysis by neighbor-joining (MEGA6).
We coded large insertions as 1 event by excluding all but 1 nt/insertion. Generated gaps were treated as missing data. Neighbor-joining analyses used either uncorrected distances or maximum-likelihood estimates of distances with a general time reversible model (6ST), empirical base frequencies, and either no rate variation among sites or a gamma distribution (shape parameter 0.5) of variation among sites with 4 rate categories. Support for branches was estimated as the percentage of neighbor-joining trees containing the branch on the basis of neighbor-joining analysis of maximum likelihood distances of 1,000 bootstrapped datasets.
PCR Amplification and Analysis by Using Basic Local Alignment Search Tool
Figure 1

Figure 1. Infected tissues from 6 bottlenose dolphins (Tursiops truncatus) with paracoccidioidomycosis ceti, Indian River Lagoon, Florida, USA, showing typical branching chains of yeast-like cells of Paracoccidioides brasiliensis connected by small isthmuses. A)...
Microscopically, the 6 silver-stained specimens showed branching chains of yeast-like cells connected by small isthmuses, which is typical of this pathogen from infected dolphins with lacaziosis/lobomycosis (Figure 1). PCR amplified the 151-bp DNA sequence from each of the genomic DNAs from the 6 dolphin formalin-fixed tissues. These DNA sequences were deposited into GenBank under accession nos. KX239500 for SW0704, KX239501 for FB946, KX239502 for FB921, KX239503 for FB 952, KX239504 for FB938, and KX239505 for B92-932. Primers targeting other DNA sequences >300 bp did not produce amplicons for all 6 DNA specimens.
Figure 2

Figure 2. Nucleotide sequences of partial Kex gene exons of Lacazia loboi (Ll) and Paracoccidioides brasiliensis (Pb), including pathogen DNA sequences isolated from bottlenose dolphins, Indian River Lagoon, Florida, USA, and P. lutzii...
Alignment of P. brasiliensis and L. loboi sequences from humans available in GenBank showed that partial Kex gene sequences of these fungi from dolphins were similar to those of P. brasilienis from humans. The only difference between P. brasiliensis sequences from humans and those from dolphins was a gap caused by a missing nucleotide in P. brasiliensis sequence from dolphins (Figure 2). P. lutzii and L. loboi sequences had several nucleotide mismatches and long gaps caused by several missing nucleotides (Figure 2). BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) analysis showed that the 6 partial Kex gene sequences had 100% homology with 7 P. brasiliensis sequences (GenBank accession nos. EU870193, EF672178, EF672177, EU870183, EU870177, EU870176, and EF672176), 93% homology with 5 P. lutzii sequences (GenBank accession nos. EF672176, EU870176, AF486805, EU870183, and EU870177), and 73% homology with 4 L. loboi sequences (GenBank accession nos. EU167516, EU167517, EU167518, and EU167519).
Phylogenetic Analysis
Figure 3
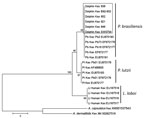
Figure 3. Unrooted maximum-parsimony phylogenetic tree of partial Kex gene sequences of Paracoccidioides brasiliensis (Pb) from 6 bottlenose dolphins, Indian River Lagoon, Florida, USA, with skin granulomas and homologous sequences of P. brasiliensis,...
Figure 4
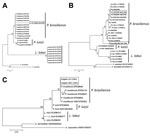
Analysis of homologous partial CHS4, Gp43, Kex, and ITS sequences of P. brasiliensis, P. lutzii, L. loboi, A. capsulatus, and A. dermatitidis (the 2 Ajellomyces species sequences were used as outgroups) by parsimony and neighbor-joining showed that dolphin-derived pathogen sequences could be placed among P. brasiliensis sequences isolated from humans with paracoccidioidomycosis (Figure 3). P. lutzii and L. loboi resolved into 2 low-supported clusters. The partial Kex gene sequences of L. loboi available in GenBank placed this uncultivated pathogen among Paracoccidioides species (Figure 3). Placement of dolphin pathogen Kex gene sequences within the cluster of P. brasiliensis was also phylogenetically corroborated by using CHS4, Gp43, and ITS sequences available in GenBank (Figure 4). Dolphin-derived pathogen sequences clustered with good bootstrap support among sequences of P. brasiliensis isolates from humans.
We found that fungal DNA sequences isolated from dolphins with skin granulomas containing yeast-like cells had strong homology with sequences of cultivated P. brasiliesis from humans (14–16). Since cutaneous granulomas containing chains of yeast-like cells in 3 dolphin species (Sotalia guainensis, T. aduncus, and T. truncatus) were initially reported, the etiologic agent of lacaziosis/lobomycosis was believed to be L. loboi, which causes similar skin granulomas in humans (1–7,19). This hypothesis was based on phenotypic characteristics of the pathogen (uniform size yeast-like cells in chains connected by slender isthmuses and resistance to culture) and clinical presentation (keloidal-like granulomas) in humans and dolphins with lacaziosis/lobomycosis (1,4,19). Although some authors had reported minor phenotypic differences, such as smaller size of yeast-like cells in infected dolphins than of yeast-like cells in infected humans (20), the true phenotypic differences between the causative agent of keloidal-like skin infections in dolphins and humans are not fully understood (4,20).
Studies using serum samples from humans and dolphins with lacaziosis/lobomycosis, mice experimentally infected with L. loboi, and serum samples from humans with paracocidioidomycosis showed that IgG in serum samples from dolphins and humans infected with L. loboi had strong cross-reactivity with the gp43 antigen of P. brasiliensis (4,6,21). These findings support the hypothesis that the uncultivated organism causing cutaneous granulomas in humans and dolphins was L. loboi. Findings also implied that the gp43 antigen of the etiologic agent of parakeloidal-like granulomas in humans and dolphins was antigenically similar to that of P. brasiliensis. On the basis of these serologic studies (4,6,21), current phylogenetic data for gp43 and Kex gene exons, and ITS DNA sequences, placement of L. loboi from humans in its own genus is questionable. Efforts to culture the organism from dolphins on classical laboratory media successfully used to isolate P. brasiliensis from humans with paracoccidioidomycosis were not successful (4,7). The physiologic basis of the inability to culture the etiologic agent from dolphins with cutaneous granulomas is not known. Thus, the life cycle features of this agent remain an enigma.
Our phylogenetic (parsimony) analysis of partial Kex DNA sequences validated reports suggesting that keloidal-like lesions in dolphins are caused by a novel uncultivated P. brasiliensis (13–16). We analyzed DNA sequences of pathogens isolated from 6 dolphins with lacaziosis/lobomycosis captured in the Indian River Lagoon. Diverse geographic locations of dolphins in the Atlantic Ocean (13,16) and the Pacific Ocean (14,15) and specimens evaluated by molecular methods provide additional support for placement of the etiologic agent of keloidal-like granulomas in dolphins within P. brasiliensis (Figure 3). Because these geographic locations, especially for cases from Japan (14,15), have different ecologic niches than locations for P. brasiliensis in South America (4), detection of dolphins infected with an uncultivated P. brasiliensis type in these ecosystems is a major finding.
Moreover, our phylogenetic data obtained by using gp43 gene exons of Minakawa et al. (14) and Ueda et al. (15), ITS sequences of Esperón et al. (16), and 2 CHS4 gene sequences (A. Schaefer, P. McCarthy, unpub. data) strongly support placement of the dolphin pathogen within cultivated P. brasiliensis isolates from humans (Figure 4). The distance between ITS sequences from dolphins and P. brasiliensis ITS sequences from humans is large (Figure 4, panel A). An evaluation of additional ITS sequences from dolphin uncultivated P. brasiliensis strains from dolphins is needed to determine if this variation indicates 2 different populations or rapid substitutions in this DNA region.
Molecular data for dolphins in the Pacific and Atlantic Oceans in previous studies (13–16), the 6 pathogen DNA sequences isolated from dolphins (this study), and 2 CHS4 gene sequences (A. Schaefer, P. McCarthy, unpub. data) place the uncultivated pathogen within cultivated P. brasiliensis strains. These studies added support to the notion that a novel uncultivated P. brasiliensis, which is different from the cultivated P. brasiliensis causing human paracoccidioidomycosis and L. loboi causing parakeloidal-like lesions in humans, is the causative agent of lacaziosis/lobomycosis in dolphins. Placement of L. loboi in a different cluster from dolphin-derived uncultivated P. brasiliensis indicates that, although both pathogens have identical phenotypes and cause similar skin lesions, they have different evolutionary paths.
Disease that shows keloidal-like granulomas in humans and dolphins has been known by several different names, such as Jorge Lobo disease (4), Lobo’s disease (3,5,22), lobomycosis (1,6,13,16,19,23,24), and lacaziosis (11,12,14,15). In view of most recent findings, the names used to describe this disease in dolphins are no longer supported. Minakawa et al. (14) proposed maintaining the name lacaziosis with the understanding that this name would include L. loboi (humans), uncultivated Paracoccidioides species, and P. brasiliensis (dolphins). However, in our phylogenetic analysis, the Paracoccidioides sp. strain (GenBank accession no. AB811031) of Ueda et al. (15) from an infected dolphin grouped among human P. brasiliensis strains. Thus, this strain is phylogenetically similar to strain LC057206. Furthermore, phylogenetic analysis of ITS sequences from dolphins with lacaziosis/lobomycosis placed L. loboi (with strong bootstrap support) in its own genus (Figure 3, panel A). Thus, the proposal by Minakawa et al. (14). could add more confusion to the taxonomic status of these 2 uncultivated fungal etiologies. In the interim, we propose paracoccidioidomycosis ceti for the disease caused by uncultivated P. brasiliensis in dolphins. This term best describes the current status of infected dolphins with keloidal-like granulomas and yeast-like cells in chains in infected tissues.
Uncultivated P. brasiliensis from Japan that causes skin infections in a new species of dolphins (Lagenorhynchus obliquidens) suggests that the geographic distribution of this pathogen is expanding and could also infect other species (14,15). Thus, whales and other cetaceans need to be investigated for this pathogen (14). Paniz-Mondolfi et al. (24) suggested that distinguishing apparent expansions of the ecologic niche caused by increased interest and surveillance by identification programs from a change in distribution would be difficult. The likelihood that this phenomenon is an expansion of its ecologic niche caused by global climate changes or increased surveillance is difficult to prove, but it is an intriguing possibility.
Although the ITS sequences of L. loboi from humans still group this pathogen in its own cluster, our molecular data for DNA protein-coding sequences indicate that the 3 species in this study (cultivated and uncultivated P. brasiliensis, P. lutzii from humans and dolphins, and L. loboi from humans) all have the same ancestor. Thus, all 3 species belong to the same genus (Paracocidioides). Comprehensive phylogenetic and genomic analyses of L. loboi from humans and uncultivated P. brasiliensis from dolphins are needed to corroborate results of these analyses and identify the true evolutionary history of L. loboi from humans. Our findings could stimulate new interest in lacaziosis and paracoccidioidomycosis ceti, which has been restricted to humans in South America and dolphins in many oceans.
Dr. Vilela is a physician in the Faculty of Pharmacy, Federal University of Minas Gerais, Belo Horizonte, Brazil. Her research interests include Lacazia loboi, Lagenidium spp., Pythium insidiosum, and Rhinosporidium seeberi.
Acknowledgments
We thank Wayne McFee for performing age analysis on wild dolphins. Free-ranging dolphins were collected under National Marine Fisheries Service Scientific Research permit no. 998-1678 to G.D.B. as part of the Bottlenose Dolphin Health and Risk Assessment Project conducted in the Indian River Lagoon, Florida, and the estuarine waters of Charleston, South Carolina.
This study was supported in part by the Department of Microbiology and Molecular Genetics, Michigan State University.
References
- Bossart GD. Suspected acquired immunodeficiency in an Atlantic bottlenosed dolphin with chronic-active hepatitis and lobomycosis. J Am Vet Med Assoc. 1984;185:1413–4.PubMedGoogle Scholar
- Bossart GD, Schaefer AM, McCulloch S, Goldstein J, Fair PA, Reif JS. Mucocutaneous lesions in free-ranging Atlantic bottlenose dolphins Tursiops truncatus from the southeastern USA. Dis Aquat Organ. 2015;115:175–84.DOIPubMedGoogle Scholar
- deVries GA, Laarman JJ. A case of Lobo’s disease in the dolphin Sotalia guianesis. Journal of Aquatic Mammals. 1973;1:26–33.
- da Lacaz S, Baruzzi RG, Rosa CB. Lobo’s disease [in Portuguese]. São Paulo: Editora da Universidad de IPSIS Gráfica e Editora; 1986.
- Migaki G, Valerio MG, Irvine B, Garner FM. Lobo’s disease in an atlantic bottle-nosed dolphin. J Am Vet Med Assoc. 1971;159:578–82.PubMedGoogle Scholar
- Woodard JC. Electron microscopic study of lobomycosis (Loboa loboi). Lab Invest. 1972;27:606–12.PubMedGoogle Scholar
- Schaefer AM, Reif JS, Guzmán EA, Bossart GD, Ottuso P, Snyder J, et al. Toward the identification, characterization and experimental culture of Lacazia loboi from Atlantic bottlenose dolphin (Tursiops truncatus). Med Mycol. 2016;54:659–65.DOIPubMedGoogle Scholar
- Silva ME, Kaplan W, Miranda JL. Antigenic relationships between Paracoccidioides loboi and other pathogenic fungi determined by immunofluorescence. Mycopathol Mycol Appl. 1968;36:97–106.DOIPubMedGoogle Scholar
- Taborda PR, Taborda VA, McGinnis MR. Lacazia loboi gen. nov., comb. nov., the etiologic agent of lobomycosis. J Clin Microbiol. 1999;37:2031–3.PubMedGoogle Scholar
- Herr RA, Tarcha EJ, Taborda PR, Taylor JW, Ajello L, Mendoza L. Phylogenetic analysis of Lacazia loboi places this previously uncharacterized pathogen within the dimorphic Onygenales. J Clin Microbiol. 2001;39:309–14.DOIPubMedGoogle Scholar
- Vilela R, Mendoza L, Rosa PS, Belone AF, Madeira S, Opromolla DV, et al. Molecular model for studying the uncultivated fungal pathogen Lacazia loboi. J Clin Microbiol. 2005;43:3657–61.DOIPubMedGoogle Scholar
- Vilela R, Rosa PS, Belone AFF, Taylor JW, Diório SM, Mendoza L. Molecular phylogeny of animal pathogen Lacazia loboi inferred from rDNA and DNA coding sequences. Mycol Res. 2009;113:851–7.DOIPubMedGoogle Scholar
- Rotstein DS, Burdett LG, McLellan W, Schwacke L, Rowles T, Terio KA, et al. Lobomycosis in offshore bottlenose dolphins (Tursiops truncatus), North Carolina. Emerg Infect Dis. 2009;15:588–90.DOIPubMedGoogle Scholar
- Minakawa T, Ueda K, Tanaka M, Tanaka N, Kuwamura M, Izawa T, et al. Detection of multiple budding yeast cells and a partial sequence of 43-kDa glycoprotein coding gene of Paracoccidioides brasiliensis from a case of lacaziosis in a female Pacific white-sided dolphin (Lagenorhynchus obliquidens). Mycopathologia. 2016;181:523–9.DOIPubMedGoogle Scholar
- Ueda K, Sano A, Yamate J, Nakagawa EI, Kuwamura M, Izawa T, et al. Two cases of lacaziosis in bottlenose dolphins (Tursiops truncates) in Japan. Case Reports in Veterinary Medicines. 2013;2013 [cited 2016 Sep 2]. https://www.hindawi.com/journals/crivem/2013/318548/
- Esperón F, García-Párraga D, Bellière EN, Sánchez-Vizcaíno JM. Molecular diagnosis of lobomycosis-like disease in a bottlenose dolphin in captivity. Med Mycol. 2012;50:106–9.DOIPubMedGoogle Scholar
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–80.DOIPubMedGoogle Scholar
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–9.DOIPubMedGoogle Scholar
- Reif JS, Mazzoil M, McCulloch SD, Varela RA, Goldstein JD, Fair PA, et al. Lobomycosis in Atlantic bottlenose dolphins (Tursiops truncatus) from the Indian River Lagoon, Florida. J Am Vet Med Assoc. 2006;228:104–8.DOIPubMedGoogle Scholar
- Haubold EM, Cooper CR Jr, Wen JW, McGinnis MR, Cowan DF. Comparative morphology of Lacazia loboi (syn. Loboa loboi) in dolphins and humans. Med Mycol. 2000;38:9–14.DOIPubMedGoogle Scholar
- Mendoza L, Belone AFF, Vilela R, Rehtanz M, Bossart GD, Reif JS, et al. Use of sera from humans and dolphins with lacaziosis and sera from experimentally infected mice for Western Blot analyses of Lacazia loboi antigens. Clin Vaccine Immunol. 2008;15:164–7.DOIPubMedGoogle Scholar
- Symmers WS. A possible case of Lôbo’s disease acquired in Europe from a bottle-nosed dolphin (Tursiops truncatus). Bull Soc Pathol Exot Filiales. 1983;76:777–84.PubMedGoogle Scholar
- Kiszka J, Van Bressem M-F, Pusineri C. Lobomycosis-like disease and other skin conditions in Indo-Pacific bottlenose dolphins Tursiops aduncus from the Indian Ocean. Dis Aquat Organ. 2009;84:151–7.DOIPubMedGoogle Scholar
- Paniz-Mondolfi A, Talhari C, Sander Hoffmann L, Connor DL, Talhari S, Bermudez-Villapol L, et al. Lobomycosis: an emerging disease in humans and delphinidae. Mycoses. 2012;55:298–309.DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 22, Number 12—December 2016
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Leonel Mendoza, Biomedical Laboratory Diagnostics Program, Department of Microbiology and Molecular Genetics, Michigan State University, North Kedzie Hall, 354 Farm Lane, Rm 324, East Lansing, MI 48824-1031, USA
Top