Volume 22, Number 9—September 2016
Research
Staphylococcus aureus Regulatory RNAs as Potential Biomarkers for Bloodstream Infections
Cite This Article
Citation for Media
Abstract
Staphylococcus aureus is a commensal bacterium and pathogen. Identifying biomarkers for the transition from colonization to disease caused by this organism would be useful. Several S. aureus small RNAs (sRNAs) regulate virulence. We investigated presence and expression of 8 sRNAs in 83 S. aureus strains from 42 patients with sepsis or septic shock and 41 asymptomatic colonized carriers. Small pathogenicity island sRNAs sprB and sprC were clade specific. Six sRNAs had variable expression not correlated with clinical status. Expression of RNAIII was lower in strains from septic shock patients than in strains from colonized patients. When RNAIII was associated with expression of sprD, colonizing strains could be discriminated from strains in patients with bloodstream infections, including patients with sepsis and septic shock. Isolates associated with colonization might have sRNAs with target expression different from those of disease isolates. Monitoring expression of RNAIII and sprD could help determine severity of bloodstream infections.
Staphylococcus aureus causes many community-acquired, healthcare–related, and nosocomial infections in humans. This bacterium is a commensal organism (part of the normal microflora), but it can also infect the body at various sites. Diseases caused by S. aureus differ greatly, ranging from skin lesions to invasive infections. A total of 20%–30% of the healthy population is colonized with S. aureus in the nostrils (1), and a substantial percentage of S. aureus bacteremia originates from endogenous colonies in the nasal mucosa (2,3).
Clinical expression of sepsis covers a continuum of manifestations; the most serious form is septic shock. In this state, vascular offense and systemic inflammation lead to endangered cardiac function and decrease in blood pressure that cause impaired oxygen delivery, organ failure, and death. Sepsis-related deaths and lack of mitigating clinical approaches attest to our limited understanding of the complex host–S. aureus interactions.
S. aureus has high rates of transmission and increased levels of antimicrobial drug resistance and produces many virulence factors (4). To coordinate expression of virulence genes during infection, S. aureus uses 2-component systems, transcription factors (5) and regulatory or small RNAs (sRNAs), which function as positive (6) or negative (7) virulence determinants. There are ≈160 sRNAs in the Staphylococcal Regulatory RNA database (8). Although their functions have not been extensively investigated, some sRNAs are known to regulate virulence factors. Quorum sensing is mediated by the accessory gene regulator agr, and RNAIII is the effector (9). Staphylococcal infection severity is based on host factors and bacterial pathogenesis (10). We investigated differences in sRNA gene content and expression levels in S. aureus strains isolated from patients with bloodstream infections and from asymptomatic carriers.
Isolates and Sample Collection
We obtained clinical isolates from a prospective study of all patients given a diagnosis of S. aureus bloodstream infections in 2006 at the Rennes University Hospital (Rennes, France), a tertiary referral hospital in western France. We selected patients with nonsevere sepsis or septic shock (11). Patients with severe sepsis (sepsis with organ dysfunction or tissue hypoperfusion improving after fluid therapy and not requiring vasopressors) were not included because their clinical status might too closely resemble nonsevere sepsis or shock. To prevent other confounding factors, immunodeficient patients were excluded: those infected with HIV; those with congenital immunodeficiency, malignant hemopathy, organ or stem cell transplants; those receiving systemic corticosteroid therapy for >3 weeks; and those undergoing another immunosuppressive treatment.
We extracted data from medical records. A nosocomial bloodstream infection was defined as either a bloodstream infection diagnosed in a patient hospitalized for >48 hours before symptom onset or a bloodstream infection in a patient receiving chronic hemodialysis or peritoneal dialysis. For each patient, we calculated at admission the Charlson Comorbidity Index and the Simplified Acute Physiology Score (12). We also collected 41 isolates from asymptomatic carriers: 23 from medical students in Rennes; 7 from healthcare workers sampled during their medical visit at a hospital in Lausanne, Switzerland; and 11 from the National Reference Laboratory for Staphylococci in Lyon, France. The study was approved by review board at Rennes University Hospital.
Multilocus Sequence Typing and S. aureus Protein A Typing
S. aureus protein A (spa) typing was performed by using primers spa-1113f (5′-TAAAGACGATCCTTCGGTGAGC-3′) and spa-1514r (5′CAGCAGTAGTGCCGTTTGCTT-3′). Sequences were determined by using a BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) and a 3730xl DNA analyzer (Applied Biosystems). The spa repeats and types were determined by using BioNumerics (Applied Maths, Sint-Matens-Latem, Belgium) and the Ridom Spa Server (http://www.spaserver.ridom.de/). The spa types with similar profiles were grouped within similar lineages. Multilocus sequence typing (MLST) was performed according to procedures of Enright et al. (13). The PCR products were sequenced by using a 3730xl DNA analyzer, and sequence types (STs) were determined by using BioNumerics and the MLST database (http://www.mlst.net/). MecA1 and MecA2 primers were used to amplify a 1,102-bp gene fragment, which was used to identify mecA. Isolates were screened for tst and pvl by using real-time PCR. PCR was used to detect small pathogenicity island (PI) RNAs (spr); sprA1/2, sprB, sprC, sprD, sprX, ssrA, 6S RNA, and rsaE. Primers used are listed in Technical Appendix Table). All PCR products were analyzed by using 2% agarose gel electrophoresis.
Bacterial Cultures, RNA Isolation, and Expression Analysis
S. aureus strains were grown in Luria-Bertani medium and then harvested. Cells were isolated by centrifugation and dissolved in a solution of 33 mmol/L sodium acetate, 17 mmol/L sodium dodecyl sulfate, and 1 mmol/L EDTA (pH 5.5). The cells were then mixed with glass beads and lysed by using a Fast Prep Apparatus (MP Biochemicals, LLC, Santa Ana, CA, USA).
RNAs were isolated by using water-saturated phenol (pH 5.0). RNAs were precipitated and washed with ethanol. Northern blotting of RNA markers was conducted by loading 10 μg of total RNA onto 8 mol/L urea, 8% polyacrylamide gels. Gels were subjected to electrophoresis and blotted onto nylon membranes at 30 V for 1.5 h in 0.5× Tris-HCl, borate, EDTA buffer.
Prehybridization and hybridization were performed by using ExpressHyb solution (Clontech, Mountain View, CA, USA) and 32P-labeled DNAs (Technical Appendix Table 2). Signals were detected by using phosphorimaging (Biocompare, South San Francisco, CA, USA) and quantified. Expression levels of sRNAs in strains were monitored by using quantitative PCR and specific primers (Technical Appendix Table 3). cDNAs were produced by using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Using the comparative cycle threshold method (Applied Biosystems), we normalized sRNA counts against transfer–messenger RNA (tmRNA) and S. aureus reference strain L102.
Bacterial Protein Extracts and Western Blotting
For preparation of protein extracts, bacteria were grown until the desired optical density (OD) at 600 nm was reached. Cells were centrifuged at 8,000 × g for 10 min at 4°C and suspended in lysis buffer (10 mmol/L Tris-HCl [pH 7.5], 20 mmol/L NaCl, 1 mmol/L EDTA, and 5 mmol/L MgCl2) in the presence of a protease inhibitor cocktail tablet containing 0.1 mg/mL lysostaphin. The cells was then dissolved in 1× Laemmli buffer containing 10% β-mercaptoethanol and heated at 90°C for 5 min. Samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis on 8% polyacrylamide gels and transferred to polyvinylidene fluoride membranes at 100 V for 1 h. Membranes were blocked in Tris-buffered saline containing 5% milk.
Second immunoglobulin-binding protein (Sbi), which is an immune evasion protein, was detected by using specific antibodies as described (14), and SaeR protein was detected by using specific antibodies. Incubation with primary antibodies against Sbi (diluted 1:10,000) or antibodies against SaeR protein (diluted 1:5,000) was performed at room temperature for 2 h. Blots were incubated with antirabbit IgG peroxidase-conjugated secondary antibodies for 1 h; washed in Tris-buffered saline, 0.05% Tween; developed in ECL Western Blotting Detection Reagent (GE Healthcare Bio-Sciences, Pittsburgh, PA, USA); and exposed on an ImageQuant LAS4000 Imaging System (GE Healthcare Bio-Sciences). Quantifications were performed by using the ImageQuant System. Levels of Sbi or SaeR protein were normalized against levels of total proteins.
All statistical tests and graphic representations were performed by using GraphPad Prism software (GraphPad Software Inc., La Jolla, CA, USA). Quantitative values were compared by using the Mann-Whitney U test. A p value <0.05 was considered significant.
Characteristics of Patients with S. aureus Bloodstream Infections
A total of 42 patients (17 with septic shock and 25 with nonsevere sepsis) were included in this study (Table 1). Patients with nonsevere sepsis were more likely than those with septic shock to have nosocomial bloodstream infections (p = 0.02) and a lower Simplified Acute Physiology Score (p = 0.01). The mortality rate was also significantly higher for patients with septic shock than for patients with nonsevere sepsis (41.2% vs. 8.0%; p = 0.01). The clonal distribution of isolates (Table 2) was similar to that reported for France in the European Antimicrobial Resistance Surveillance System (15).
Genotyping of Strains from Patients with Invasive Diseases and Asymptomatic Carriers
Figure 1
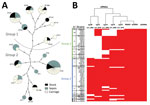
Figure 1. Phylogenetic and molecular typing of Staphylococcus aureus by using small RNAs (SRNAs), Rennes, France. A) Phylogenetic tree showing sRNA gene content and expression. Genotyping by multilocus sequence typing (MLST) and the...
We used MLST and spa typing to analyze 83 S. aureus isolates from 42 blood cultures for patients with bloodstream infections or 41 nasal samples from asymptomatic carriers. Isolates clustered into 17 STs (Figure 1, panel A). Of 83 strains analyzed, none had genes encoding the Panton-Valentine leukocidin, which is associated with increased virulence of certain strains. Toxic shock syndrome toxin genes were detected in infectious and methicillin-susceptible S. aureus colonizing strains that belonged to ST5 and ST30.
Among 37 methicillin-susceptible S. aureus colonizing strains, ST398 was the most common (n = 6) (Technical Appendix Table 4). Only 4 strains isolated from nasal samples were methicillin-resistant S. aureus (MRSA), and these belonged to ST8 and ST22 (Table 2). Most (n = 10) MRSA isolates were ST8 (Table 2). MRSA prevalence was ≈10% in healthy colonized healthcare workers and students and ≈2% in the general population (17). The prevalence of MRSA in infectious samples (≈21%), all positive for mecA, was consistent with the overall prevalence of staphylococcal infections in France (18). We detected a predominance of the ST8 MRSA clone, which is the predominant pandemic MRSA clone in France and has sea and lukED genes. Isolates from patients with bloodstream infections and carriers were evenly distributed among the STs (Table 2). As reported by Feil et al. (19), genetic distances between the 8 group 1 STs (mean ± SD 25.2 ± 10.6) were longer than those between the 9 group 2 STs (20.0 ± 7.4), which is consistent with earlier emergence of group 1 isolates.
Specificities of sRNAs for Bacterial Clades
We used PCR to identify a subset of sRNAs that were specific for conserved sequences. Eight sRNAs were selected from core and accessory genomes for determining their distribution among strains. We chose sRNAs according to their presence in the accessory genome because this presence implies variability in their presence/absence among strains and their putative roles in virulence. We included the few sRNAs ubiquitously detected in bacteria. Housekeeping tmRNA and 6S RNA genes, which were detected in many bacterial species, were found in all strains. All strains also contained RNAIII, which is the quorum-sensing effector (9).
In addition, we detected 5 sRNAs expressed on PIs: sprA (srn_3580), sprB (srn_3600), sprC (srn_3610), sprD (srn_3800), and sprX (srn_3820) (8). Because of absence of PIs phiSa3 and vsaβ in group 1, all 5 sRNAs on PIs were detected only in group 2 STs (Table 2). We found that sprA was rarely detected in ST398 strains, sprB was not detected in all group 1 strains and STs, sprC was not detected in group 1 STs except ST398 strains, sprD was detected in all but 5 strains from both groups, and sprX was detected in all strains except ST398. Detection of sprD and sprX in most STs from both groups reflects evolution of S. aureus, which has been punctuated by successive acquisitions and losses of genetic elements. The presence of sprB and sprC among S. aureus infectious isolates indicates S. aureus phylogeny and strain clonality (Figure 1, panel B). Strain genotyping showed that the sample analyzed reflected diversity of staphylococcal infections at the national level in France.
PI-Encoded RNA Expression
Figure 2

Figure 2. Expression of 5 sRNAs and tmRNA in 16 Staphylococcus aureus isolates from patients with bloodstream infections (nonsevere sepsis or septic shock) and asymptomatic colonized carriers (carriage), Rennes, France. A) sprA; B)...
Figure 3

Figure 3. Expression of sprD in 61 Staphylococcus aureus isolates, Rennes, France. A) Representative growth curves. Each curve indicates 1 strain. Arrows indicate times at which the total RNAs were collected. E, early...
Because of low amounts (10–100 CFU/mL of blood) of bacteria isolated from patients with bloodstream infections (20), S. aureus isolates must be cultured before assessing sRNA expression. We selected 16 strains for subsequent analyses: 5 from nasal carriers, 6 from patients with sepsis, and 5 from patients with septic shock. Each group of samples contained strains from the same sequence types (ST5, ST8, and ST25) (Table 2). We intentionally included strains belonging to the same ST (ST8), with strains from nasal carriers, from patients with sepsis, and from patients with septic shock. Strain selection was also dictated by availability in our collection. For these 16 isolates, we assessed sRNA expression levels at OD600 nm = 2 (early exponential), OD600 nm = 4 (late exponential), and OD600 nm = 8 (stationary) growth phases (Figure 2; Technical Appendix Figure). Growth curves for all isolates were superimposable (Figure 3, panel A).
We compared overall sRNA expression levels among the infectious subgroups. For all strains, tmRNA was constitutively expressed, and there were no differences in expression among strains (Figure 2). This finding is consistent with the status of the tmRNA gene as a housekeeping gene involved in ribosome rescue (21). The 6S RNA was also constitutively expressed, and there were no differences in expression among strains. Expression of the 5 Spr RNAs varied widely among the strains (Figure 2). We found that sRNA showed different expression profiles within the same ST (Technical Appendix Figure), which indicated the complexity and variability of sRNA-driven gene regulation in S. aureus.
Expression of sprD was heterogeneous in strains from asymptomatic carriers but more homogeneous in infected patients (Figure 2). These results were inferred from Northern blotting performed for 3 independent RNA extractions. Afterwards, the set of analyzed strains was nearly quadrupled to 61 isolates, with 21 from carriers, 23 from nonsevere sepsis patients, and 17 from septic shock patients. Because Northern blotting showed variations in SprD expression levels between the clinical strain sets (Figure 2), we monitored SprD expression by using quantitative PCR at OD600nm = 2 (Figure 3, panel A). Strains from asymptomatic carriers and sepsis patients expressed SprD heterogeneously, although SprD was expressed at low levels in all strains isolated from patients with septic shock (Figure 3, panel B).
Discrimination of Isolates by Expression Levels of RNAIII and RNAIII/SprD
Figure 4
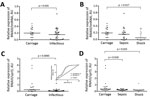
Figure 4. Discrimination of colonizing strains of Staphylococcus aureus from patients with bloodstream infections, Rennes, France. A) RNAIII analysis of strains from carriers and infected persons; B) RNAIII analysis of strains from carriers...
Another RNA implicated in S. aureus virulence is RNAIII (9), which is an archetype of RNA-mediated regulation of virulence genes. Therefore, we used quantitative PCR to monitor RNAIII expression levels for 61 isolates (Figure 4) during E growth phase (Figure 3, panel A). We detected significantly lower RNAIII levels in strains isolated from patients with bloodstream infections than in those from nasal carriers (p = 0.035) (Figure 4, panel A). When we compared strains from patients with septic shock with commensal isolates, strains from patients with septic shock had significantly lower levels of RNAIII (p = 0.017) (Figure 4, panel B). Calculated RNAIII expression levels in infectious and asymptomatic persons showed a progressive decrease from carriage to nonsevere sepsis to septic shock. When we combined the effects of SprD with RNAIII, this combination substantially discriminated carriage isolates from infectious isolates (p = 0.0065) (Figure 4, panel C), carriage isolates from sepsis isolates (p = 0.018), and carriage isolates from septic shock isolates (p = 0.025) (Figure 4, panel D). We conducted receiver operating characteristic analyses to determine the capacity of differential expression of RNAIII and SprD in predicting disease outcome. Our findings indicate differences in RNAIII/SprD expression levels between colonizing strains and infectious strains (Figure 4, panel C, inset).
Discrimination of Isolates by Expression Levels of Sbi
Figure 5
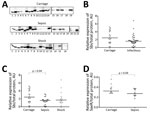
Figure 5. Expression levels of Sbi and SaeR proteins of Staphylococcus aureus isolated from patients with bloodstream infections and asymptomatic carriers, Rennes, France. A) Western blot of Sbi protein for 61 strains during...
SprD and RNAIII negatively regulate expression of Sbi by blocking translation through binding with the Sbi mRNA (6,14). We performed Western blotting for 61 isolates in growth phase E by using polyclonal antibodies against intracellular and membrane proteins to monitor Sbi levels (Figure 5, panel A). As reported by Smith et al. (22), we found that molecular weights of Sbi detected in isolates were variable (mean ≈50 kDa) and the amount of Sbi varied among isolates (Figure 5, panel A). Individual assessments showed significantly lower Sbi levels in isolates from patients with bloodstream infections than in patients with nasal colonization (p = 0.04).
A set of 83 S. aureus strains with known genotypes was collected from asymptomatic carriers and patients with nonsevere sepsis or septic shock. We used this collection for a prospective study of the presence and expression of certain sRNAs in the core and accessory genome. We also monitored expression of Sbi, an immune evasion protein whose expression is negatively controlled by the sRNAs SprD and RNAIII (14) and SaeR, a positive regulator of Sbi. In clinical and carriage staphylococcal strains, the presence of >2 PI-encoded sRNAs, sprB (srn_3600) and sprC (srn_3610), was indicative of the presence of PIs and prophages. These PI-encoded sRNAs, particularly sprB, could be used as probes to improve genotyping studies. These sRNAs probably appear during the transition between ST22 and ST25 (Figure 1, panel A). The sRNA sprB is absent in most group 1 isolates. In some strains, we cannot rule out that sequence variations among the sRNA genes might hamper their amplification.
Molecular typing can identify genetic diversity of strains, which is required for epidemiologic surveillance of infections. Bacterial strain typing methods include generation of DNA banding patterns, sequencing, and hybridization. Bacterial genotyping has benefited from identification of novel locus-specific typing markers. The sRNAs might be useful probes for genotyping bacteria because their overall content varies considerably, even among closely-related strains. Furthermore, because several sRNAs are encoded by mobile genetic elements, these sRNAs reflect the acquisition/loss of virulence factors encoded by these elements and molecules that confer antimicrobial drug resistance. In phylogenetic studies, selected sRNAs located in accessory genomes might shed light on genomic diversity.
The immunologic ability of patients to eradicate pathogens is a major determinant of infection outcome, and patients with septic shock are often immunocompromised (23). Our data suggests that, for staphylococcal bloodstream infections, one must also consider attributes of infecting strains, including at least an immune evasion molecule and sRNAs RNAIII and sprD. The effector of the agr quorum-sensing system was expressed at lower levels in strains isolated from patients with bloodstream infections, especially those with septic shock, than in asymptomatic carriers. Therefore, low RNAIII levels might identify S. aureus isolates that are responsible for bloodstream infections, even after isolation and culture. agr-defective S. aureus is a frequent cause of bloodstream infections (24). Our observations concur with the previous identification of inactivating mutations in the S. aureus agr virulence regulator, which have been associated with poor outcomes in for patients with bloodstream infections (3). Coupling the expression levels of RNAIII and SprD could discriminate colonization from infection and also provide useful information about severity of bloodstream infections. However, this possibility must be confirmed with a larger set of clinical isolates.
Because sprD and RNAIII negatively control the expression of the Sbi immune evasion protein (17), we also monitored Sbi expression in the isolates. Sbi is an immune evasion factor (25) involved in the S. aureus-induced inflammatory response (26). Cell wall−anchored Sbi proteins are essential components for S. aureus survival in the commensal state (27). Detection of additional Sbi proteins in strains isolated from asymptomatic carriers than in those from patients with septic shock is consistent with their role during colonization and in immune tolerance. Expression of Sbi at the transcriptional level is positively regulated by SaeRS, which is composed of the histidine kinase SaeS and the response regulator SaeR (28). Expression of Sbi is controlled by >3 regulators: negatively by 2 sRNAs, and positively by a 2-component system. This system indicates why RNAIII and sprD levels are not inversely correlated with Sbi levels in isolates tested. Carriage strains have higher SaeR levels than the sepsis strains, which is consistent with higher Sbi levels in the carriage strains than sepsis strains (Figure 5).
The transition from commensalism to infection in S. aureus is an essential but complex process. From a clinical standpoint, most S. aureus infections are derived from previous colonizers (29). When those strains switch to invasiveness, the transition may be related to regulatory network expression changes, including within sRNAs. Sequencing has identified changes in regulatory functions of strains isolated from a person who initially was a carrier and then showed development of a fatal bloodstream infection (30), which suggested that molecular evolution might play a key role in this process. Our results suggest that certain sRNAs from the gene regulatory network in a human pathogen will provide insights into commensal-to-pathogen transitions. These sRNAs could be used as surrogate markers for severity of staphylococcal infections and as biomarkers for prophylaxis and monitoring of S. aureus infection. Comparison of frequency and expression of selected sRNAs in S. aureus isolates that colonize or become infectious might be a way to identify associations between sRNA expression and disease patterns.
In vitro expression levels of some S. aureus sRNAs might not reflect their in vivo levels (31). Nevertheless, direct analysis of expression levels of bacterial sRNAs directly in blood from patients with bloodstream infections is technically difficult because of low levels of bacteria collected. We compared sRNA expression in fresh isolates from patients with the same isolates after they were thawed after be frozen for 3 weeks. We found no differences in RNAIII and sprD expression between isolates obtained directly from patients and isolates that had been frozen. Subsequent investigations will address the functional and clinical relevance of expression patterns of RNAIII and sprD. Broadening our investigations to include additional sRNAs might identify biomarkers that predict staphylococcal disease severity in infected patients. In addition to their roles as biomarkers, sRNAs could also be targets for innovative therapeutic approaches.
Dr. Bordeau is a research engineer at University of Rennes 1, Rennes, France. Her primary research interests are functions and structures of sRNAs.
Acknowledgments
We thank T. Bae for providing antibodies against SaeR protein; F. Laurent, M. Bes, and O. Sakwinska for providing strains from asymptomatic carriers; and C. Daniela for providing assistance with statistical analysis.
A patent for the new RNA biomarkers reported in this article has been filed by the Institut National de la Santé et de la Recherche Médicale Transfert (Paris, France).
This study was supported by the Institut National de la Santé et de la Recherche Médicale (grant to B.F.). M.S. was supported by the Region Bretagne (grants SAD SARS #8254 and SAD SARS_2 #9181).
References
- van Belkum A, Verkaik NJ, de Vogel CP, Boelens HA, Verveer J, Nouwen JL, Reclassification of Staphylococcus aureus nasal carriage types. J Infect Dis. 2009;199:1820–6 .DOIPubMedGoogle Scholar
- von Eiff C, Becker K, Machka K, Stammer H, Peters G. Nasal carriage as a source of Staphylococcus aureus bacteremia. Study Group. N Engl J Med. 2001;344:11–6 .DOIPubMedGoogle Scholar
- Smyth DS, Kafer JM, Wasserman GA, Velickovic L, Mathema B, Holzman RS, Nasal carriage as a source of agr-defective Staphylococcus aureus bacteremia. J Infect Dis. 2012;206:1168–77 .DOIPubMedGoogle Scholar
- Tong SY, Davis JS, Eichenberger E, Holland TL, Fowler VG Jr. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28:603–61 .DOIPubMedGoogle Scholar
- Rosenstein R, Gotz F. What distinguishes highly pathogenic staphylococci from medium- and non-pathogenic? Curr Top Microbiol Immunol. 2013;358:33–89 .DOIPubMedGoogle Scholar
- Chabelskaya S, Gaillot O, Felden B. A Staphylococcus aureus small RNA is required for bacterial virulence and regulates the expression of an immune-evasion molecule. PLoS Pathog. 2010;6:e1000927 .DOIPubMedGoogle Scholar
- Romilly C, Lays C, Tomasini A, Caldelari I, Benito Y, Hammann P, A non-coding RNA promotes bacterial persistence and decreases virulence by regulating a regulator in Staphylococcus aureus. PLoS Pathog. 2014;10:e1003979 .DOIPubMedGoogle Scholar
- Sassi M, Augagneur Y, Mauro T, Ivain L, Chabelskaya S, Hallier M, SRD: a Staphylococcus regulatory RNA database. RNA. 2015;21:1005–17 .DOIPubMedGoogle Scholar
- Novick RP, Geisinger E. Quorum sensing in staphylococci. Annu Rev Genet. 2008;42:541–64 .DOIPubMedGoogle Scholar
- Powers ME, Bubeck Wardenburg J. Igniting the fire: Staphylococcus aureus virulence factors in the pathogenesis of sepsis. PLoS Pathog. 2014;10:e1003871 .DOIPubMedGoogle Scholar
- Dellinger RP, Levy MM, Rhodes A, Annane D, Gerlach H, Opal SM, Surviving sepsis campaign: international guidelines for management of severe sepsis and septic shock, 2012. Intensive Care Med. 2013;39:165–228 .DOIPubMedGoogle Scholar
- Christensen S, Johansen MB, Christiansen CF, Jensen R, Lemeshow S. Comparison of Charlson Comorbidity Index with SAPS and APACHE scores for prediction of mortality following intensive care. Clin Epidemiol. 2011;3:203–11 .DOIPubMedGoogle Scholar
- Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38:1008–15.PubMedGoogle Scholar
- Chabelskaya S, Bordeau V, Felden B. Dual RNA regulatory control of a Staphylococcus aureus virulence factor. Nucleic Acids Res. 2014;42:4847–58 .DOIPubMedGoogle Scholar
- Dauwalder O, Lina G, Durand G, Bes M, Meugnier H, Jarlier V, Epidemiology of invasive methicillin-resistant Staphylococcus aureus clones collected in France in 2006 and 2007. J Clin Microbiol. 2008;46:3454–8 .DOIPubMedGoogle Scholar
- Cooper JE, Feil EJ. The phylogeny of Staphylococcus aureus: which genes make the best intra-species markers? Microbiology. 2006;152:1297–305 .DOIPubMedGoogle Scholar
- Robert J, Tristan A, Cavalie L, Decousser JW, Bes M, Etienne J, Panton-Valentine leukocidin–positive and toxic shock syndrome toxin 1–positive methicillin-resistant Staphylococcus aureus: a French multicenter prospective study in 2008. Antimicrob Agents Chemother. 2011;55:1734–9 .DOIPubMedGoogle Scholar
- Le Moing V, Alla F, Doco-Lecompte T, Delahaye F, Piroth L, Chirouze C, Staphylococcus aureus bloodstream infection and endocarditis: a prospective cohort study. PLoS One. 2015;10:e0127385 .DOIPubMedGoogle Scholar
- Feil EJ, Cooper JE, Grundmann H, Robinson DA, Enright MC, Berendt T, How clonal is Staphylococcus aureus? J Bacteriol. 2003;185:3307–16 .DOIPubMedGoogle Scholar
- Kiehn TE, Wong B, Edwards FF, Armstrong D. Comparative recovery of bacteria and yeasts from lysis-centrifugation and a conventional blood culture system. J Clin Microbiol. 1983;18:300–4.PubMedGoogle Scholar
- Keiler KC. Mechanisms of ribosome rescue in bacteria. Nat Rev Microbiol. 2015;13:285–97 .DOIPubMedGoogle Scholar
- Smith EJ, Visai L, Kerrigan SW, Speziale P, Foster TJ. The Sbi protein is a multifunctional immune evasion factor of Staphylococcus aureus. Infect Immun. 2011;79:3801–9 .DOIPubMedGoogle Scholar
- Hotchkiss RS, Monneret G, Payen D. Immunosuppression in sepsis: a novel understanding of the disorder and a new therapeutic approach. Lancet Infect Dis. 2013;13:260–8 .DOIPubMedGoogle Scholar
- Painter KL, Krishna A, Wigneshweraraj S, Edwards AM. What role does the quorum-sensing accessory gene regulator system play during Staphylococcus aureus bacteremia? Trends Microbiol. 2014;22:676–85 .DOIPubMedGoogle Scholar
- Burman JD, Leung E, Atkins KL, O’Seaghdha MN, Lango L, Bernado P, Interaction of human complement with Sbi, a staphylococcal immunoglobulin-binding protein: indications of a novel mechanism of complement evasion by Staphylococcus aureus. J Biol Chem. 2008;283:17579–93 .DOIPubMedGoogle Scholar
- Gonzalez CD, Ledo C, Giai C, Garofalo A, Gomez MI. The Sbi protein contributes to Staphylococcus aureus inflammatory response during systemic infection. PLoS One. 2015;10:e0131879 .DOIPubMedGoogle Scholar
- Foster TJ, Geoghegan JA, Ganesh VK, Hook M. Adhesion, invasion and evasion: the many functions of the surface proteins of Staphylococcus aureus. Nat Rev Microbiol. 2014;12:49–62 .DOIPubMedGoogle Scholar
- Rogasch K, Ruhmling V, Pane-Farre J, Hoper D, Weinberg C, Fuchs S, Influence of the two-component system SaeRS on global gene expression in two different Staphylococcus aureus strains. J Bacteriol. 2006;188:7742–58 .DOIPubMedGoogle Scholar
- Wertheim HF, Melles DC, Vos MC, van Leeuwen W, van Belkum A, Verbrugh HA, The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis. 2005;5:751–62 .DOIPubMedGoogle Scholar
- Young BC, Golubchik T, Batty EM, Fung R, Larner-Svensson H, Votintseva AA, Evolutionary dynamics of Staphylococcus aureus during progression from carriage to disease. Proc Natl Acad Sci U S A. 2012;109:4550–5 .DOIPubMedGoogle Scholar
- Song J, Lays C, Vandenesch F, Benito Y, Bes M, Chu Y, The expression of small regulatory RNAs in clinical samples reflects the different life styles of Staphylococcus aureus in colonization vs. infection. PLoS One. 2012;7:e37294 .DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 22, Number 9—September 2016
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Brice Felden, Department of Pharmaceutical Biochemistry, University of Rennes 1, 2 Av Professor Léon Bernard, Rennes, Brittany 35043, France
Top