Volume 31, Number 6—June 2025
Research
Emergence of Oropouche Virus in Espírito Santo State, Brazil, 2024
Cite This Article
Citation for Media
Abstract
Oropouche virus (OROV), historically endemic to the Amazon, had spread to nearly all Brazil states by 2024; Espírito Santo emerged as a transmission hotspot in the Atlantic Forest biome. We characterized the epidemiologic factors driving OROV spread in nonendemic southeast Brazil, analyzing environmental and agricultural conditions contributing to viral transmission. We tested samples from 29,080 suspected arbovirus-infected patients quantitative reverse transcription PCR for OROV and dengue, chikungunya, Zika, and Mayaro viruses. During March‒June 2024, the state had 339 confirmed OROV cases, demonstrating successful local transmission. Spatial analysis revealed that most cases clustered in municipalities with tropical climates and intensive cacao, robusta coffee, coconut, and pepper cultivation. Phylogenetic analysis identified the Espírito Santo OROV strains as part of the 2022–2024 Amazon lineage. The rapid spread of OROV outside the Amazon highlights its adaptive potential and public health threat, emphasizing the need for enhanced surveillance and targeted control measures.
Oropouche virus (OROV), classified as Orthobunyavirus oropoucheense, family Peribunyaviridae, is an arthropodborne virus with a negative-sense RNA genome consisting of 3 segments, large (L), medium (M), and small (S) (1). A neglected arbovirus, OROV causes Oropouche fever and circulates primarily in Central America, South America, and the Caribbean (2–4). In Brazil, OROV was historically confined to the Amazon basin, where several vector species and a range of reservoirs maintain its sylvatic transmission cycle (2,3,5). In urban areas in the Amazon, Culicoides paraensis, a midge commonly found in tropical, humid environments rich in organic matter, such as forests and plantations, is the primary vector responsible for OROV transmission to humans (3,4). Humans might acquire OROV infection in forested regions and subsequently introduce it to urban settings. The widespread distribution of the vector, coupled with increased human mobility and the influence of climate change, might enable the virus’s gradual expansion beyond its historical range in Brazil, raising concerns about the potential for broader geographic spread (6,7).
Since the 1960s, occasional OROV spillovers to humans have led to >30 documented localized outbreaks or large-scale epidemics in the Amazon basin, underscoring the virus’s epidemic potential (8–12). Although incidence is highest in the Amazon, sporadic cases have been reported in other states in Brazil without leading to widespread outbreaks (13).
During August 2022‒March 2024, a new outbreak triggered by a reassortant OROV lineage emerged in Brazil’s western Amazon region, causing ≈6,000 reported cases (14). In 2024, that reassortant lineage led to the largest recorded outbreak outside the virus’s endemic zone; OROV was detected in all regions of Brazil (15,16). Outside the Amazon, high incidence rates were observed in the Atlantic Forest region, particularly in municipalities with low population densities and agricultural activities favoring the establishment of C. paraensis vector populations, such as cocoa and banana cultivation (17).
OROV infections typically manifest as an acute febrile illness characterized by headache, myalgia, and arthralgia (14). Those symptoms overlap with those of infection with other endemic arboviruses, such as dengue virus (DENV), Zika virus (ZIKV), and chikungunya virus (CHIKV) (2,3). However, emerging evidence has linked OROV to fatal cases (18). Unprecedented vertical transmission was also reported, and some cases resulted in congenital anomalies or fetal death (19,20). Therefore, the significant shift in OROV’s pathogenicity marks a new epidemiologic paradigm for Oropouche fever.
Espírito Santo state, located in southeastern Brazil and entirely within the Atlantic Forest biome, has emerged as a major hotspot for OROV transmission outside the Amazon region, recording the highest state-level incidence rate among non-Amazon states in 2024 (21). The state’s extensive agricultural activities, particularly in coffee, cocoa, and banana cultivation (22), combined with high rural worker mobility and environmental conditions favorable for C. paraensis midge establishment, may have contributed to viral spread. The first case of Oropouche infection in Espírito Santo was detected on March 24, 2024. We used epidemiologic and genomic approaches to analyze the emergence and dissemination of the new OROV variant in Espírito Santo state. We also examined the regional characteristics that enable its transmission and contribute to its establishment in this previously unaffected area.
Study Population
Our study analyzed samples from patients who visited public health units in Espírito Santo state with arboviral-like symptoms (ZDC: Zika, dengue, and chikungunya) during February 25–June 15, 2024. All suspected cases of acute arboviral infections in Espírito Santo are centralized at the state’s central laboratory, Laboratório Central de Saúde Pública do Espírito Santo (LACEN-ES, Vitória, Brazil), which receives samples from all 78 municipalities for diagnostic testing. During the study period, a total of 29,080 samples were tested, corresponding to ≈0.76% of the state’s population (3,833,712 inhabitants). The serum or plasma samples were subsequently sent to the LACEN-ES for viral molecular diagnosis. The Human Research Ethics Committee at the University of Vila Velha (Vila Velha, Brazil) approved this study (Certificate of Presentation for Ethical Assessment no. 84698324.7.0000.5064) and waived the need for written informed consent. The committee is registered with Brazil’s National Research Ethics Commission and oversees studies involving public health institutions in Espírito Santo, including the state’s Central Laboratory where this investigation was conducted.
Sample Processing and Quantitative Reverse Transcription PCR
We processed samples for nucleic acid extraction using magnetic bead-based systems TANBead Maelstrom 9600 (Taiwan Advanced Nanotech Inc., https://www.tanbead.com), EXTRACTA 96 (Loccus, https://www.loccus.com.br) and TechStar YC-702 (Wuxi Techstar Technology Co., http://www.techstarbio.com), following manufacturers’ instructions. Subsequently, for molecular testing, we used 3 different quantitative reverse transcription (qRT-PCR) kits: Molecular ZCD Tipagem Bio-Manguinhos (https://www.bio.fiocruz.br), Biomol ZDC (IBMP, https://www.ibmp.org.br), and VIASURE Zika, Dengue & Chikungunya (ThermoFisher, https://www.thermofisher.com), following the manufacturers’ protocols. For the samples not detectable for ZDC, we performed a multiplex qRT-PCR to investigate for OROV and Mayaro virus (MAYV), as described by Naveca et al. (23). To extend testing, we adapted the laboratory diagnosis to carry out the procedure on a pool of 8 samples. After detecting the target, we conducted another q RT-PCR with the 8 samples individually to confirm the test.
Epidemiologic Data and Environmental Context
We retrieved individual-level of OROV-positive case data from the eSUS Brazil Ministry of Health (https://sisaps.saude.gov.br/sistemas/esusaps) and Gerenciador de Ambiente Laboratorial (GAL; http://gal.datasus.gov.br) systems at the municipal level for the state of Espírito Santo, covering cases reported through June 15, 2024. We linked each case to anonymized metadata, including demographics, location, symptoms (fever, headache, myalgia, retroorbital pain, back pain, arthritis, petechiae, rash, arthralgia, nausea, conjunctivitis, vomiting, and leukopenia), and hospitalization and notification/symptom dates.
We calculated the municipal incidence of OROV on the basis of the 2022 census data from the Brazilian Institute of Geography and Statistics (IBGE) (24). Estimated incidence mapping used the geoBR package in RStudio (http://www.rstudio.com) with IBGE municipal boundary shapefiles (25). We obtained data about the agricultural establishments at the municipal level from the 2017 IBGE census of agriculture (26). We estimated the Spearman correlation for all municipalities reporting cases to explore the relationship between the planted area of the top 10 crops in Espírito Santo and the number of OROV cases. In addition, we analyzed between-group differences in cycle threshold (Ct) values (viral load proxy) for each symptom using 2-tailed Mann-Whitney U tests (α = 0.05).
Generation Time and Instantaneous Reproduction Number Estimation
The generation time represents the interval between successive rounds of infection. Although that interval has been estimated for other arboviruses, no such estimates exist for OROV. We estimated the generation time using a combination of human viral clearance data (27), mosquito mortality rates (28), and data from experimental studies involving vector competence (29), using a framework applied previously for ZIKV (30) and MAYV (31). We conducted parameter inference using a Bayesian framework implemented with Markov chain Monte Carlo methods (Appendix Figures 1, 2). We then used the posterior distributions of the generation time parameters (Appendix Tables 1, 2) to inform the calculation of the reproduction number. We estimated the instantaneous reproduction number (Rt) for OROV using the R package EpiEstim (32). We reconstructed the Rt for the April–June period using a Bayesian inference model with a sliding time window of τ = 7 days (Appendix).
Sample Selection and Next-Generation Sequencing
We selected 7 samples with Ct <27 for whole-genome sequencing from a pool of all OROV-positive samples identified through RT-PCR during the study. To ensure geographic representativeness, we chose samples from the municipalities of Colatina, Rio Bananal, and Laranja da Terra, which were among the most affected regions in the state. We sequenced 6 samples at Instituto Adolfo Lutz (IAL), yielding complete S and M segments (Appendix Table 3). We sequenced 1 sample at LACEN/ES using Illumina RNA Prep with Enrichment Tagmentation protocol (https://www.illumina.com) with the Respiratory Pathogen ID/AMR Enrichment Panel Kit (Illumina); we extracted OROV reads from the nontarget portion of the kit. We conducted genome assembly using a custom version of the ViralFlow pipeline version 1.0.1 (33), referencing GenBank sequences NC_005776.1 (L segment), NC_005775.1 (M segment), and NC_005777.1 (S segment).
Bioinformatics and Phylogenetic Inference
For the phylogenetic analysis, we aligned 7 new sequences from Espírito Santo with 145 OROV strains sampled in the Americas (1955–2023) and available in GenBank as of August 2024. The alignment, performed in MAFFT version 7 (https://mafft.cbrc.jp) (34), included the prototypical viruses Iquitos, Madre de Dios, and Perdões as outgroups. We selected the generalized time-reversible discrete gamma substitution model in jModelTest2 (https://github.com/ddarriba/jmodeltest2). We used phylogenetic inference in MrBayes version 3.2.7a (35) to sample trees until parameter convergence (effective sample size >200), with node support determined by posterior probabilities from the majority-rule consensus topology.
Figure 1
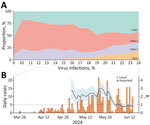
Figure 1. Temporal distribution and reproduction number of Oropouche virus in Espírito Santo, Brazil. A) Weekly percentages of CHIKV, DENV-1, DENV-2, and OROV infections for epidemiologic weeks 9–24, 2024. B) Estimates of...
Approximately 29,100 patients in Espírito Santo experiencing arbovirus-like illness were tested by real-time RT-PCR for the presence of DENV-1, DENV-2, CHIKV, ZIKV, OROV, and MAYV during March–June 2024. Until epidemiologic week 13, DENV-1 represented ≈50% of the positive cases, followed by CHIKV and DENV-2 (Figure 1). However, from epidemiologic week 13 onward, OROV cases were detected, marking a pronounced shift in the epidemiologic landscape. OROV cases increased rapidly, reaching 339 cases within 10 weeks. By epidemiologic week 24, the frequency of OROV infections approached the levels of DENV-1, DENV-2, and CHIKV, indicating a comparable circulation of these viruses at the outbreak’s peak. After the emergence of OROV, the proportion of CHIKV cases also rose, eventually surpassing that of DENV. Initially, OROV cases were primarily classified as imported; however, community spread became evident as local transmission was established. During epidemiologic weeks 18–27, the time-varying Rt for OROV remained ≈2.5 (95% credible interval ≈1.5–3.0), we observed a peak value of ≈3.0 in epidemiologic week 26. After this peak, a marked reduction in Rt occurred, and it was below 1.0 by epidemiologic week 28, indicating a decline in transmission and a shift toward containment of the outbreak (Figure 1).
Figure 2
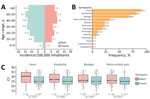
Figure 2. Demographics, symptoms, and Ct value comparison in OROV-positive patients from Espírito Santo, Brazil. A) Age-sex pyramid of the OROV incidence rate (per 100,000 inhabitants). Incidence rates are shown for each...
We observed a marked predominance of male patients in OROV cases (Figure 2, panel A); male-to-female ratio was 1.5:1. Most cases occurred in persons >20 years of age, suggesting that adult men may be disproportionately affected by this outbreak. The most frequently reported symptoms were fever (307/339 [90.56%]), headache (275/339 [81.12%]), myalgia (232/339 [68.44%]), and retroorbital pain (113/339 [33.33%]) (Figure 2, panel B). We observed no significant differences in symptoms between male and female patients. We used real-time RT-PCR Ct values as a proxy for the patient’s plasmatic viral load to investigate its relationship to symptomatology. Patients with fever exhibited significantly lower median Ct values than those without fever (p = 0.017), suggesting a higher viral load in febrile patients. We observed a similar trend for headache (p = 0.043), myalgia (p = 0.025), and retroorbital pain (p = 0.017) (Figure 2, panel C). Comparisons for other symptoms did not yield statistically significant differences (Appendix Figure 1).
Figure 3
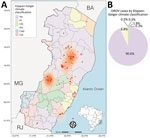
Figure 3. Spatial distribution of Oropouche virus cases and climate classification in Espírito Santo, Brazil. A) Espírito Santo, with municipalities colored according to the Köppen-Geiger climate classification. Red dots indicate cases. Orange...
During the OROV outbreak in Espírito Santo, the virus spread across 17 municipalities, culminating in 8.84 cases/100,000 inhabitants statewide. Most cases were concentrated in 2 distinct hotspots: the regions surrounding the municipalities of Colatina/Rio Bananal and Laranja da Terra (Figure 3, panel A). In total, 332/339 (98%) of the diagnosed cases were associated with tropical climates, in accordance with Köppen-Geiger climate classification; 308 (≈91%) occurred in municipalities classified as having a tropical savanna climate (Köppen-Geiger classification Aw), and 24 (7%) were linked to tropical monsoon climate (Köppen-Geiger classification Am) (Figure 3, panel B). In contrast, only 7/339 cases (≈2%) were distributed across municipalities with temperate climates, such as humid subtropical (Köppen-Geiger classification Cfa) and temperate oceanic (Köppen-Geiger classification Cfb). No cases were reported in areas classified as subtropical highland climate (Köppen-Geiger classification Cwb).
Figure 4
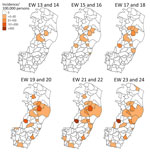
Figure 4. Oropouche virus case incidence by municipality and 2-week epidemiologic period, Espírito Santo state, Brazil. EW, epidemiologic week.
The initial spread of OROV in Espírito Santo followed a clear spatial pattern, predominantly affecting municipalities within the tropical savanna climate (Figure 4). This dissemination phase, epidemiologic weeks 17–18, marked the entry of the virus into areas with different climatic conditions. The peak incidence, exceeding 200 cases/100,000 inhabitants, was recorded at the municipality level during epidemiologic weeks 21–24, particularly in the epicentral municipalities of Colatina and Laranja da Terra. Those areas, situated within the tropical savanna climate zone, appear to have served as primary foci for transmitting OROV throughout the state. To characterize the ecologic niches contributing to the introduction and spread of OROV in Espírito Santo, we examined the association between OROV prevalence and the cultivated areas for the 10 most widely grown crops in the state. We found significant associations between specific crops and OROV incidence (Table); robusta coffee (Spearman correlation coefficient [ρ] = 0.55, p = 0.004), cacao (ρ = 0.54, p = 0.005), coconut (ρ = 0.43, p = 0.003), and pepper (ρ = 0.43, p = 0.034) displayed moderate positive correlations with the number of OROV cases.
Figure 5

Figure 5. Bayesian phylogenetic trees created from analysis of Oropouche virus segments associated with the reassortant lineage M1L2S2in Espírito Santo, Brazil, and reference sequences. Each tree...
Phylogenetic analysis of each genomic segment individually showed that all cases detected within Espírito Santo belong to the novel reassortant M1L2S2 (OROVBR2015–2024) lineage (15) with high branch support (posterior probability >0.98). That lineage circulated in the Amazon Basin during 2023–2024, causing a major outbreak (Figure 5). Of interest, in the S and M trees, the Espírito Santo samples do not form a monophyletic clade, which indicated multiple introductions of OROV into the state.
The emergence and rapid spread of arboviruses beyond their traditional endemic regions is driven by changing climate patterns and human land use (7). Our study used high-resolution data to document the unprecedented establishment of OROV in Brazil’s Atlantic Forest, showing its adaptations and spread beyond the Amazon Basin. The expansion mirrors patterns observed for other arboviruses, such as ZIKV (36), CHIKV (37), and West Nile virus (38), in which changes in vector ecology, human mobility, and environmental conditions have led to emergence of those viruses in previously unaffected regions (6). OROV’s successful establishment in southeast Brazil underscores both an immediate public health concern and the complex ecologic and epidemiologic factors enabling arboviral adaptation to novel environments.
Our findings revealed a marked epidemiologic shift with the emergence and rapid establishment of OROV transmission in the state amidst the ongoing endemic circulation of DENV and CHIKV. Within ≈11 weeks from its initial detection, OROV reached infection levels comparable to those of endemic arboviruses in Espírito Santo, suggesting robust viral transmission. The concurrent presence of OROV alongside DENV and CHIKV, without evident competitive interference, could reflect vector-specific ecologic differences: although DENV and CHIKV are transmitted predominantly by Aedes spp. mosquitoes in urban areas (39), OROV transmission is driven mainly by C. paraensis midges (3,27). The estimated transmissibility, with Rt peaking at ≈3.0, parallels the dynamics seen in both urban and sylvatic arboviruses within immunologically naive or mixed populations. Similar patterns were reported for ZIKV; Rt values were 2.6–4.8 in French Polynesia (40), 3.0–6.6 in Colombia (41), and as high as 2.5 in Brazil (30). Likewise, MAYV, endemic to the Amazon region, showed Rt values of 2.1–2.9 within its primary range, decreasing to 1.1–1.3 in non-Amazon regions (31). The high transmissibility of OROV observed in Espírito Santo could be attributed to multiple factors, including the predominantly mild or asymptomatic nature of OROV infections (3,27) and favorable local conditions for C. paraensis midge proliferation (28). Our Rt estimates might underestimate transmissibility by excluding unreported asymptomatic and mild cases. However, if underreporting remains stable, the inferred trends should still reliably reflect epidemic dynamics (32).
The demographic profile of affected persons in this outbreak showed predominance of adult male case-patients, which contrasts with previous OROV outbreaks (12,14). The male predominance may reflect occupational exposure in rural agricultural activities, increasing contact with C. paraensis vectors, as seen in previous Brazil outbreaks where farming was a major risk factor (42). Clinical manifestations aligned with previous OROV cases across diverse regions, including recent Amazon outbreaks (12,14,43). The association between key symptoms and lower Ct values suggested that higher viral loads may drive the intensity of clinical manifestations, as documented for other arboviruses, such as DENV (44,45), yet remains largely unexplored for OROV. Of note, recent fatal OROV cases in nonendemic regions of Brazil, characterized by low Ct values and rapid progression to severe hemorrhagic symptoms within days, underscored the potential link between viral load and disease severity (18).
Phylogenetic analysis of OROV genomic sequences from Espírito Santo revealed a complex pattern of viral spread, with multiple independent introductions of the virus into the state. The samples clustered within the OROVBrazil2015–2024 clade, specifically within the M1L2S2 reassortant lineage that caused the recent outbreak in the Amazon region (14). The absence of monophyly in the S and M segment trees suggests that multiple introduction events occurred followed by local spread through different routes, as described in Gräf et al. (17). That molecular pattern corroborates the epidemiologic data showing the initial emergence of imported cases followed by rapid establishment of autochthonous transmission in different municipalities. Further genomic surveillance is needed to fully understand the dispersal patterns and the state’s role in the virus’s spread to other regions, highlighting the need for comprehensive and continuous surveillance throughout the region.
The predominance of OROV transmission in regions with tropical climates (Köppen-Geiger classifications Aw and Am) highlights the critical role of environmental conditions that favor a high prevalence of potential vectors, particularly C. paraensis midges. Although comprehensive studies on Culicoides spp. distribution across climate zones of Brazil are lacking, studies in Europe have documented strong associations between Köppen climate classifications and Culicoides spp. diversity (46). The tropical climates of Espírito Santo outbreak zones have high temperatures and seasonal rainfall, likely creating optimal conditions for C. paraensis breeding. Those environments provide abundant organic matter for larval development; decomposing vegetation, fruit waste, banana tree stumps, and cacao husks, common agricultural byproducts in the region, are ideal breeding sites (3,4,27). Indeed, studies report peak C. paraensis midge populations during the rainy season; temperatures are 30°C–32°C and relative humidity 75%–85% (47), favorable macroclimatic and microclimatic conditions for vector proliferation. In contrast, temperate and subtropical highland climates, characterized by lower temperatures, more evenly distributed rainfall, and distinct vegetation, present less favorable conditions for vector establishment. Despite the widespread distribution of Culicoides spp. midges across the Americas (28), no cases of OROV had been reported in Espírito Santo state until 2024. The introduction of OROV into Espírito Santo is estimated to have occurred multiple times during February–March 2024 (17), and its establishment may reflect ecologic shifts that have amplified vector density or increased human–vector contact, particularly among susceptible populations in Espírito Santo and other regions of Brazil. Espírito Santo’s recent biting midge infestation in 2019 (48) likely supported conditions for OROV emergence by increasing vector populations in previously unaffected areas. Climate change projections in Europe suggest a shift toward warmer and wetter conditions that may transform humid subtropical climates into subtropical climates with hot summers, further promoting Culicoides spp. establishment (46). Modeling studies indicate that climate-driven increases in temperature and extreme weather events could expand arboviral transmission in tropical regions like Espírito Santo, as observed for DENV (49). These ecologic changes likely interact with agricultural and land-use patterns, shaping OROV transmission and creating corridors that intensify virus-vector-host interactions. Further research is warranted to quantify the relationships between climatic variables, C. paraensis species dynamics, and OROV transmission in Espírito Santo and other countries.
The significant correlations between OROV incidence and specific crops in Espírito Santo underscore the complex interactions between agricultural landscapes and arboviral transmission dynamics. Outside the Amazon region, OROV transmission has been predominantly associated with rural settings, where C. paraensis midges find optimal breeding conditions. C. paraensis larvae develop effectively in microhabitats created by decaying organic matter from banana and cacao plantations (3,4,27); of note, recent outbreaks outside the Amazon have occurred along the Atlantic Forest biome where these crops are prevalent (17). The emergence of OROV in Espírito Santo’s regions with high densities of robusta coffee, pepper, and coconut cultivation represents a novel association that warrants further investigation, because it would broaden our understanding of potential agricultural landscapes that may support C. paraensis populations. That pattern aligns with the vector’s documented ability to occupy both sylvatic and anthropic environments (28) and suggests that agricultural expansion may create ecologic pathways that encourage viral spread. The fragmentation of forest landscapes by agricultural activities likely intensifies human–vector contact, particularly in areas where multiple crops create diverse microhabitats suitable for vector breeding. The strong presence of OROV in Espírito Santo’s major agricultural regions, especially where coffee and cacao cultivation predominate, emphasizes the need for detailed ecologic studies of C. paraensis midges in those settings to identify and predict potential outbreak hotspots.
Our study provides valuable insights into environmental and ecologic factors influencing OROV emergence in Espírito Santo. A limitation is that inferences about spatial dissemination relied on assumptions regarding the primary vector, C. paraensis midges, without directly assessing its competence, density, or dispersal potential, which may affect estimates of transmission dynamics. In addition, reliance on secondary epidemiologic data, rather than targeted field sampling, limits the granularity and completeness of information on transmission patterns and hotspots. Observed correlations between OROV cases and crops might also reflect geographic clustering effects, potentially introducing spatial biases. Future research with fine-scale spatial modeling and systematic vector surveillance is essential to clarify interactions among environmental variables, vector ecology, and OROV transmission dynamics.
In conclusion, our study reveals a substantial epidemiologic shift marked by the emergence and establishment of OROV in Espírito Santo, underscoring the virus’s capacity to adapt to new ecologic landscapes outside its Amazon origins. The rapid rise in OROV cases to levels comparable with established arboviruses like DENV and CHIKV, without apparent competitive inhibition, highlights the distinct ecologic niches exploited by Culicoides spp. vectors in periurban and rural areas. The interplay between tropical climate, expanding agricultural landscapes, and favorable breeding sites for C. paraensis midges likely enabled this outbreak, positioning Espírito Santo as a potential hotspot for arboviral transmission amid environmental and climatic changes. As such, our findings call for heightened surveillance of both human cases and vector populations along with further investigation into the eco-epidemiologic drivers of OROV in Atlantic Forest regions. Understanding those dynamics will be crucial to predict, prevent, and respond to future outbreaks of OROV and other emerging arboviruses in similar environments.
Dr. Delatorre is a professor and researcher at the Federal University of Espírito Santo, specializing in viral genomics and infectious disease epidemiology. His research focuses on the evolutionary dynamics and spatial-temporal dispersion of emerging viruses, with an emphasis on genomic surveillance and public health responses.
Acknowledgments
We thank the staff at Laboratório Central de Saúde Pública do Espírito Santo and the Adolfo Lutz Institute for their invaluable contributions to this study. Their dedication to public health was instrumental in the successful completion of our research and in advancing scientific knowledge in this field. We also thank Gabriel Lessa Lavagnoli for his assistance in implementing the framework for generation time estimation.
All viral sequence data generated in this study are available in GenBank (accession nos. PQ197397–405).
This study was partially funded by a grant from the State of Espírito Santo Government through the Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (FAPES) (grant no. 2025-25N6P-DI 006/2025-SESA/SEAG/FAPES). This study also received institutional support from the Brazilian Ministry of Health and the Espírito Santo State Government. A.R. is a doctoral fellow funded by FAPES under the PROCAP 2025 program (grant no. 2025-0Z48T, DI 106/2025). F.G.N. is supported by FAPEAM call 04/2022/FIOCRUZ/FAPEAM/FAPERO–Inovação na Amazônia; FAPEAM–call 023/2022–Iniciativa Amazônia+10; and Conselho Nacional de Desenvolvimento Científico e Tecnológico–CNPq–Chamada CNPq/MCTI 10/2023– Faixa B–Grupos Consolidados–Universal 2023 (421620/2023-4). The authors declare no conflicts of interest.
This article was preprinted at https://www.medrxiv.org/content/10.1101/2024.12.11.24318883v1.full.pdf.
References
- de Souza WM, Calisher CH, Carrera JP, Hughes HR, Nunes MRT, Russell B, et al. ICTV virus taxonomy profile: Peribunyaviridae 2024. J Gen Virol. 2024;105:
002034 . DOIPubMedGoogle Scholar - Sakkas H, Bozidis P, Franks A, Papadopoulou C. Oropouche fever: a review. Viruses. 2018;10:175. DOIPubMedGoogle Scholar
- Travassos da Rosa JF, de Souza WM, Pinheiro FP, Figueiredo ML, Cardoso JF, Acrani GO, et al. Oropouche virus: clinical, epidemiological, and molecular aspects of a neglected orthobunyavirus. Am J Trop Med Hyg. 2017;96:1019–30. DOIPubMedGoogle Scholar
- Wesselmann KM, Postigo-Hidalgo I, Pezzi L, de Oliveira-Filho EF, Fischer C, de Lamballerie X, et al. Emergence of Oropouche fever in Latin America: a narrative review. Lancet Infect Dis. 2024;24:e439–52. DOIPubMedGoogle Scholar
- Pereira-Silva JW, Ríos-Velásquez CM, Lima GRD, Marialva Dos Santos EF, Belchior HCM, Luz SLB, et al. Distribution and diversity of mosquitoes and Oropouche-like virus infection rates in an Amazonian rural settlement. PLoS One. 2021;16:
e0246932 . DOIPubMedGoogle Scholar - de Thoisy B, Gräf T, Mansur DS, Delfraro A, Dos Santos CND. The risk of virus emergence in South America: a subtle balance between increasingly favorable conditions and a protective environment. Annu Rev Virol. 2024;11:43–65. DOIPubMedGoogle Scholar
- Baker RE, Mahmud AS, Miller IF, Rajeev M, Rasambainarivo F, Rice BL, et al. Infectious disease in an era of global change. Nat Rev Microbiol. 2022;20:193–205. DOIPubMedGoogle Scholar
- Naveca FG, Nascimento VA, Souza VC, de Figueiredo RMP. Human Orthobunyavirus Infections, Tefé, Amazonas, Brazil. PLoS Curr. 2018;10:
ecurrents.outbreaks.7d65e5eb6ef75664da68905c5582f7f7 .PubMedGoogle Scholar - Vasconcelos HB, Azevedo RSS, Casseb SM, Nunes-Neto JP, Chiang JO, Cantuária PC, et al. Oropouche fever epidemic in Northern Brazil: epidemiology and molecular characterization of isolates. J Clin Virol. 2009;44:129–33. DOIPubMedGoogle Scholar
- Bernardes-Terzian AC, de-Moraes-Bronzoni RV, Drumond BP, Da Silva-Nunes M, da-Silva NS, Urbano-Ferreira M, et al. Sporadic oropouche virus infection, acre, Brazil. Emerg Infect Dis. 2009;15:348–50. DOIPubMedGoogle Scholar
- Mourãão MP, Bastos MS, Gimaqu JBL, Mota BR, Souza GS, Grimmer GHN, et al. Oropouche fever outbreak, Manaus, Brazil, 2007-2008. Emerg Infect Dis. 2009;15:2063–4. DOIPubMedGoogle Scholar
- Azevedo RS, Nunes MRT, Chiang JO, Bensabath G, Vasconcelos HB, Pinto AYDN, et al. Reemergence of Oropouche fever, northern Brazil. Emerg Infect Dis. 2007;13:912–5. DOIPubMedGoogle Scholar
- Pan American Health Organization, World Health Organization. Epidemiological alert. Oropouche in the Region of the Americas: vertical transmission event under investigation in Brazil—July 17, 2024. 2024 [cited 2025 Apr 17]. https://www.paho.org/pt/documentos/alerta-epidemiologico-oropouche-na-regiao-das-americas-evento-transmissao-vertical-sob
- Naveca FG, Almeida TAP, Souza V, Nascimento V, Silva D, Nascimento F, et al. Human outbreaks of a novel reassortant Oropouche virus in the Brazilian Amazon region. Nat Med. 2024;30:3509–21. DOIPubMedGoogle Scholar
- Scachetti GC, Forato J, Claro IM, Hua X, Salgado BB, Vieira A, et al. Re-emergence of Oropouche virus between 2023 and 2024 in Brazil: an observational epidemiological study. Lancet Infect Dis. 2025;25:166–75. DOIPubMedGoogle Scholar
- Moreira FRR, Dutra JVR, de Carvalho AHB, Reis CR, Rios JSH, Ribeiro MO, et al. Oropouche virus genomic surveillance in Brazil. Lancet Infect Dis. 2024;24:e664–6. DOIPubMedGoogle Scholar
- Gräf T, Delatorre E, do Nascimento Ferreira C, Rossi A, Santos HGG, Pizzato BR, et al.; OROV Study Group. Expansion of Oropouche virus in non-endemic Brazilian regions: analysis of genomic characterisation and ecological drivers. Lancet Infect Dis. 2025;25:379–89. DOIPubMedGoogle Scholar
- Bandeira AC, Pereira FM, Leal A, Santos SPO, Barbosa AC, Souza MSPL, et al. Fatal Oropouche virus infections in nonendemic region, Brazil, 2024. Emerg Infect Dis. 2024;30:2370–4. DOIPubMedGoogle Scholar
- das Neves Martins FE, Chiang JO, Nunes BTD, Ribeiro BFR, Martins LC, Casseb LMN, et al. Newborns with microcephaly in Brazil and potential vertical transmission of Oropouche virus: a case series. Lancet Infect Dis. 2025;25:155–65. DOIPubMedGoogle Scholar
- Garcia Filho C, Lima Neto AS, Maia AMPC, da Silva LOR, Cavalcante RDC, Monteiro HDS, et al. A case of vertical transmission of Oropouche virus in Brazil. N Engl J Med. 2024;391:2055–7. DOIPubMedGoogle Scholar
- Secretaria de Vigilância em Saúde e Ambiente, Ministério da Saúde. Monitoring of arboviruses and closing balance of the Emergency Operations Committee (COE) dengue and other arboviruses 2024 [in Portuguese]. Boletim Epidemiológico vol. 55, no. 11. 2024 [cited 2025 Apr 17]. https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/boletins/epidemiologicos/edicoes/2024/boletim-epidemiologico-volume-55-no-11.pdf/view
- Galeano EAV, Padovan MP, de Figueiredo MRP, Takemoto ACK, Maioli HRO. Performance of agricultural production in Espírito Santo from 2010 to 2022 [in Portuguese]. Vitoria, Brazil: Incaper (Instituto Capixaba de Pesquisa, Assistência Técnica e Extensão Rural); 2024 [cited 2025 Apr 17]. https://biblioteca.incaper.es.gov.br/digital/bitstream/item/4703/1/Doc311-DesempenhodaProducadoES-Incaper.pdf
- Naveca FG, Nascimento VAD, Souza VC, Nunes BTD, Rodrigues DSG, Vasconcelos PFDC. Multiplexed reverse transcription real-time polymerase chain reaction for simultaneous detection of Mayaro, Oropouche, and Oropouche-like viruses. Mem Inst Oswaldo Cruz. 2017;112:510–3. DOIPubMedGoogle Scholar
- Instituto Brasileiro de Geografia e Estatística. 2022 Census. 2025 [cited 2025 Apr 17]. https://www.ibge.gov.br/en/statistics/social/labor/22836-2022-census-3.html
- Instituto Brasileiro de Geografia e Estatística. Geosciences downloads. 2025 [cited 2025 Apr 17]. https://www.ibge.gov.br/en/geosciences/downloads-geosciences.html
- Instituto Brasileiro de Geografia e Estatística. Census of agriculture 2017. 2025 [cited 2025 Apr 17]. https://censoagro2017.ibge.gov.br
- Pinheiro FP, Travassos da Rosa AP, Travassos da Rosa JF, Ishak R, Freitas RB, Gomes ML, et al. Oropouche virus. I. A review of clinical, epidemiological, and ecological findings. Am J Trop Med Hyg. 1981;30:149–60. DOIPubMedGoogle Scholar
- Mellor PS, Boorman J, Baylis M. Culicoides biting midges: their role as arbovirus vectors. Annu Rev Entomol. 2000;45:307–40. DOIPubMedGoogle Scholar
- McGregor BL, Connelly CR, Kenney JL. Infection, dissemination, and transmission potential of North American Culex quinquefasciatus, Culex tarsalis, and Culicoides sonorensis for Oropouche virus. Viruses. 2021;13:226. DOIPubMedGoogle Scholar
- Ferguson NM, Cucunubá ZM, Dorigatti I, Nedjati-Gilani GL, Donnelly CA, Basáñez MG, et al. EPIDEMIOLOGY. Countering the Zika epidemic in Latin America. Science. 2016;353:353–4. DOIPubMedGoogle Scholar
- Caicedo EY, Charniga K, Rueda A, Dorigatti I, Mendez Y, Hamlet A, et al. The epidemiology of Mayaro virus in the Americas: A systematic review and key parameter estimates for outbreak modelling. PLoS Negl Trop Dis. 2021;15:
e0009418 . DOIPubMedGoogle Scholar - Cori A, Ferguson NM, Fraser C, Cauchemez S. A new framework and software to estimate time-varying reproduction numbers during epidemics. Am J Epidemiol. 2013;178:1505–12. DOIPubMedGoogle Scholar
- da Silva AF, da Silva Neto AM, Aksenen CF, Jeronimo PMC, Dezordi FZ, Almeida SP, et al. ViralFlow v1.0-a computational workflow for streamlining viral genomic surveillance. NAR Genom Bioinform. 2024;6:
lqae056 . DOIPubMedGoogle Scholar - Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30:772–80. DOIPubMedGoogle Scholar
- Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012;61:539–42. DOIPubMedGoogle Scholar
- Metsky HC, Matranga CB, Wohl S, Schaffner SF, Freije CA, Winnicki SM, et al. Zika virus evolution and spread in the Americas. Nature. 2017;546:411–5. DOIPubMedGoogle Scholar
- Souza TML, Vieira YR, Delatorre E, Barbosa-Lima G, Luiz RLF, Vizzoni A, et al. Emergence of the East-Central-South-African genotype of Chikungunya virus in Brazil and the city of Rio de Janeiro may have occurred years before surveillance detection. Sci Rep. 2019;9:2760. DOIPubMedGoogle Scholar
- Kramer LD, Ciota AT, Kilpatrick AM. Introduction, Spread, and Establishment of West Nile Virus in the Americas. J Med Entomol. 2019;56:1448–55. DOIPubMedGoogle Scholar
- Segura NA, Muñoz AL, Losada-Barragán M, Torres O, Rodríguez AK, Rangel H, et al. Minireview: Epidemiological impact of arboviral diseases in Latin American countries, arbovirus-vector interactions and control strategies. Pathog Dis. 2021;79:
ftab043 . DOIPubMedGoogle Scholar - Kucharski AJ, Funk S, Eggo RM, Mallet HP, Edmunds WJ, Nilles EJ. Transmission dynamics of Zika virus in island populations: a modelling analysis of the 2013–14 French Polynesia outbreak. PLoS Negl Trop Dis. 2016;10:
e0004726 . DOIPubMedGoogle Scholar - Nishiura H, Mizumoto K, Villamil-Gómez WE, Rodríguez-Morales AJ. Preliminary estimation of the basic reproduction number of Zika virus infection during Colombia epidemic, 2015-2016. Travel Med Infect Dis. 2016;14:274–6. DOIPubMedGoogle Scholar
- Carvalho VL, Azevedo RSS, Carvalho VL, Azevedo RS, Henriques DF, Cruz ACR, et al. Arbovirus outbreak in a rural region of the Brazilian Amazon. J Clin Virol. 2022;150-151:
105155 . DOIPubMedGoogle Scholar - Silva-Caso W, Aguilar-Luis MA, Palomares-Reyes C, Mazulis F, Weilg C, Del Valle LJ, et al. First outbreak of Oropouche Fever reported in a non-endemic western region of the Peruvian Amazon: Molecular diagnosis and clinical characteristics. Int J Infect Dis. 2019;83:139–44. DOIPubMedGoogle Scholar
- Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, Suntayakorn S, et al. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis. 2000;181:2–9. DOIPubMedGoogle Scholar
- Bhatt P, Jayaram A, Varma M, Mukhopadhyay C. Kinetics of dengue viremia and its association with disease severity: an ambispective study. Virusdisease. 2024;35:250–9. DOIPubMedGoogle Scholar
- Brugger K, Rubel F. Characterizing the species composition of European Culicoides vectors by means of the Köppen-Geiger climate classification. Parasit Vectors. 2013;6:333. DOIPubMedGoogle Scholar
- Feitoza LHM, de Carvalho LPC, da Silva LR, Meireles ACA, Rios FGF, Silva GS, et al. Influence of meteorological and seasonal parameters on the activity of Culicoides paraensis (Diptera: Ceratopogonidae), an annoying anthropophilic biting midge and putative vector of Oropouche Virus in Rondônia, Brazilian Amazon. Acta Trop. 2023;243:
106928 . DOIPubMedGoogle Scholar - Prefeitura de Marechal F. Anchieta, Alfredo Chaves and Marechal unite to try to fight the advance of the maruim. 2024 [cited 2025 Apr 17]. https://www.marechalfloriano.es.gov.br/anchieta-alfredo-chaves-e-marechal-se-unem-para-tentar-combater-avanco-do-maruim
- Barcellos Madeira Rosa Y, Tamanini Silva Moschen H, Loss AC, Cardoso da Silva TC, Brioschi Dos Santos AP, Caetano Pimenta B, et al. Climate change impacts on dengue transmission areas in Espírito Santo state, Brazil. Oxf Open Immunol. 2024;5:
iqae011 . DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: May 21, 2025
Table of Contents – Volume 31, Number 6—June 2025
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Edson Delatorre, Laboratório de Genômica e Ecologia Viral, Centro de Ciências da Saúde, Universidade Federal do Espírito Santo, Av. Marechal Campos, 1468–Maruípe, 29.040-090, Vitória, ES, Brazil
Top