Volume 32, Number 7—July 2026
Online Report
Inconsistent Strategies to Mitigate the Effects of Batrachochytrium salamandrivorans, Europe
Cite This Article
Citation for Media
Abstract
Emerging infectious diseases are one of the biggest challenges in a globalized world. To date, resources have been allocated to prevent and control the spread of zoonotic and livestock pathogens. We argue that, in line with the One Health approach, equitable efforts, financial resources, attention, and coordination are required for wildlife-only pathogens to halt biodiversity loss. Deploying the amphibian fungus Batrachochytrium salamandrivorans as a model, we demonstrate the unbalanced efforts among countries in Europe regarding surveillance, disease response, prevention, public outreach, and research. We compare investments with B. salamandrivorans–free countries such as the United States, concluding that structural resources are urgently needed to curb the effects of this fungus within Europe and beyond. We encourage dialogue among authorities, researchers, and stakeholders and propose a coordinated European Union–level program of €6–10 million over 5–7 years to implement B. salamandrivorans action plans and define structural funding requirements for future wildlife disease mitigation.
The effects of the Anthropocene on ecosystem, human, and animal health renders emerging infectious diseases (EIDs) key threats in the globally recognized One Health approach. Although some EIDs transmitted from wildlife affect human or domestic animal health, others drive wildlife declines only (1–4). Those wildlife-only EIDs can have far-reaching consequences for ecosystem functioning and human health (4–7). In detail, disease-driven alterations to community composition can have cascading effects across trophic levels, resulting in major downstream impacts on nutrient cycles and food web members (1,8–12). However, despite the overall definition and interpretation of One Health, wildlife-only EIDs are not given the same priority for mitigation as zoonotic or livestock EIDs (13,14). Conversely, approaches to consider and implement wildlife diseases into One Health do exist (15,16). In this article, we urge that those approaches be expanded to wildlife-only diseases and that the mitigation of disease threats to biodiversity be established as a key component of One Health strategies (17). Using an amphibian epidemic currently observed in Europe as an example, we demonstrate that structural resources are necessary to globally curb disease-driven loss of biodiversity and propose a coordinated European Union (EU)–level funding benchmark over >5 years to support this effort and guide dialogue among authorities, stakeholders, and researchers in Europe. Their effects on biodiversity demand that EIDs are approached using integrated and collaborative efforts between different authorities (i.e., in Europe: Environment and Health and Food Safety Departments of the European Commission) and should be properly addressed within One Health.
Figure 1

Figure 1. Efforts by country in the European Union and United Kingdom in study of inconsistent strategies to mitigate the effects of Batrachochytrium salamandrivorans, Europe. A) Known B. salamandrivorans...
The skin fungal disease chytridiomycosis, induced by Batrachochytrium dendrobatidis, has caused undeniable harm to global amphibian diversity, placing it among the top 5 threats to amphibians (18,19). Similarly, another batrachochytrid fungus, Batrachochytrium salamandrivorans, is emerging, potentially threatening salamander species (Caudata) and amphibian communities outside its native range (20–24) and possibly rising as a major species conservation challenge in the upcoming decade. B. salamandrivorans, originating from Asia and likely transported to Europe by the pet trade, was discovered in 2013 in the Netherlands and can cause lethal infection in most European salamander species (20). Its recent discovery and relatively limited observed distribution suggest B. salamandrivorans to be in a relatively early stage of invasion in Europe, which would make it a prime candidate for pathogen containment and elimination (24–27). However, after 2 decades of presence in Europe, >130 known outbreaks of B. salamandrivorans chytridiomycosis have occurred in the wild, affecting >6 native species in 4 countries in Europe (Figure 1, panel A) (27,28). Although natural B. salamandrivorans dispersal appears to be a relatively slow process with an estimated range expansion of 5–16 km/year (27), long-distance and likely human-mediated dispersal and independent introductions have been documented (26–29). In addition, anurans might serve as transmission and dispersal vectors, and B. salamandrivorans has been detected in captive amphibians (caudates and anurans) in private and zoo collections in 5 countries in Europe (Figure 1, panel A) (30). The pathogen is likely to be more widespread than currently recognized because it can be carried by some host species without causing disease, and when disease does occur, clinical signs are not pathognomonic (29).
The EU initially responded to the novel threat of B. salamandrivorans by funding the development of the Batrachochytrium salamandrivorans Action Plan in 2017 (24). B. salamandrivorans has been listed in the European Animal Health Law (AHL) (31) since April 1, 2021. Of the 64 infectious diseases listed in the AHL, B. salamandrivorans–induced chytridiomycosis is the single wildlife-only EID that is subject to EU-wide regulations and the sole pathogen listed for biodiversity conservation reasons. The AHL listing of B. salamandrivorans in categories D and E dictates EU member states to conduct surveillance and prevent the spread of this pathogen. In an ideal scenario, such efforts would include both passive and active surveillance through early warning systems and targeted research, in addition to mitigation actions to curb B. salamandrivorans spread. Yet, current surveillance efforts are insufficient to detect B. salamandrivorans in its invasive range. The current observed distribution, the number of outbreaks, and the overall impact on species in Europe do not fit the predicted scenarios based on the potential host range and presumed unrestricted dispersal across most of Europe (23–29,32–34) (Figure 1, panel A). Therefore, the pathogen in Europe is still likely to be overlooked in the wild (29,33).
We compiled key information on B. salamandrivorans detection and funding until 2025 in 15 of 27 European Union countries and the United Kingdom since its discovery (Figure 1; Appendix 1 Table). We identified funding resources in 13 countries (12 EU countries and the UK) (Figure 1, panel B), as well as 16 countries with B. salamandrivorans projects (15 EU countries and the UK) (Figure 1, panel C; Appendix 2 Table). Response actions to B. salamandrivorans risk, either through funding or projects, are unknown in the other 12 countries, but are likely to be limited, if present at all. Among known actions, >60% of the tested sites and ≈71% of all samples tested for B. salamandrivorans presence (both in the wild and in captivity) were taken in the 4 countries with outbreaks in the wild (Belgium, Germany, the Netherlands, and Spain (Figure 1, panel A). Surveillance activities in natural environments and among wild amphibians occur 10 times more frequently than those targeting captive amphibians (46,578 vs. 4,940) (Appendix 1 Table), thereby limiting the capacity to detect a major transmission pathway of the disease. This discrepancy reveals either an immense Europewide sampling gap in captive amphibians, or, if testing is present but not reported, an unstructured, nontransparent reporting system for this pathogen. That deficit is further illustrated by publications with punctual data sampling (28,29,33–40).
In countries where B. salamandrivorans outbreaks have not been reported (N = 11) (Figure 1; Appendix 1 Table), only Austria and Luxembourg have sampling at a level commensurate with efforts expended in countries where the pathogen has been detected (Appendix 2 Figure). Regarding B. salamandrivorans–related funding across EU member states and the United Kingdom since the fungus’s scientific description in 2013, the overall volume reached >€12 million. So far, this money has been used for passive surveillance measures across and within member states through the establishment of a network in Europe consisting of 16 diagnostic centers and, for some countries, the development of national and regional action plans and risk analyses. These measures, combined with active measures, have led to the current knowledge on B. salamandrivorans in Europe as previously described. However, most (89%) of that funding was used by the 4 countries in which the fungus is present in the wild (Figure 1, panel A). In contrast, 7 other countries had access to funding of <€100,000 (Appendix 1 Table). Given those findings, B. salamandrivorans is likely still overlooked in Europe, and the current spatial distribution is skewed toward countries with access to funding.
Figure 2
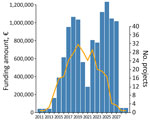
Figure 2. Overview of granted funding and projects per year in study of inconsistent strategies to mitigate the effects of Batrachochytrium salamandrivorans, Europe. Amount of funding per year is based...
Overall, 77% of the projects identified were dedicated to B. salamandrivorans only. Of the 114 B. salamandrivorans projects identified, ≈83% aimed at B. salamandrivorans in the wild, of which ≈45% had funding for >3 years. In contrast, only 7% of the projects focused on the pathogen in captive facilities only. Moreover, projects differed in their components: 66.6% of projects included the detection of the fungus and 30% included its mitigation. For projects that incorporated conservation action as a project component, 43% of projects applied in situ conservation, whereas only 6% included ex situ conservation. A combination of in situ and ex situ conservation was used in 17.5% of all projects. None included rewilding actions (Appendix 1 Table). Overall, 38.6% of all projects focused additionally or exclusively on B. salamandrivorans fundamental research with a funding volume of ≈€9.8 million (80% of total money available) (Appendix 1 Table). That investment yielded fundamental knowledge of diversity, epidemiology, and disease ecology, resulting in the development of diagnostics and treatment strategies for captive urodeles. However, additional funding is needed for continued surveillance and development of sustainable, efficient, and long-term disease mitigation of natural populations (24–27,29–35,41). Since 2013, B. salamandrivorans funding experienced 2 peaks (≈€1.1 million in 2018 and ≈€1.2 million in 2025), explained by the number of projects that started and reached their midterm in 2019 and by the number that reached their midterm and end in 2025 (Figure 2; Appendix 1 Table). So far, available (fixed) funding for 2026 and 2027 is above ≈€1 million per year followed by 2 years with funding below €100,000; that funding is shared between 4 projects only (Figure 2; Appendix 1 Table). With regard to donors, 68% (≈€8 million) came from government funding, whereas ≈3% (€417,039) were granted by nongovernmental organizations and <1% (€101,887) by universities. The EU invested ≈27% (≈€3.3 million) (Appendix 1 Table).
B. salamandrivorans sampling is insufficient in the EU for landscape-level detection and control. The limited-to-nonexistent enforcement of the AHL is exemplified by an almost complete lack of mitigation actions at known sites with infected wild populations, despite the availability of an action plan, including guidance on development of mitigation strategies (24,25). National B. salamandrivorans Action Plans exist in only 3 Member States (Belgium, Czech Republic, Luxembourg), but only Belgium and Luxembourg have incorporated the fungus into a (temporal) national strategy (Appendix 1 Table). Apart from that strategy, if actions are taken, they are mostly limited to raising public awareness and ex situ rescue of infected animals, often without a clear perspective of potential reintroduction and with uncertain effects on disease epidemiology. A few countries have invested in B. salamandrivorans research (Appendix 1 Table), but progress in both fundamental science and disease management is constrained by the paucity of studies testing the feasibility and effectiveness of mitigation or elimination strategies. Notable exceptions include a B. salamandrivorans outbreak in Catalonia, Spain, in 2018 that was met with rigorous actions that at least temporarily contained infection (26). The Netherlands has recently allocated €2 million to research on B. salamandrivorans disease ecology aiming to further develop in situ mitigation, and the EU recently invested €2.5 million (Horizon Europe/ERC101096163-GLOSSI) in fundamental research that might translate to novel B. salamandrivorans mitigation strategies in the future (e.g., long-term and in situ mitigation increasing host resistance and/or decreasing pathogen virulence). However, overall, the current, minimal efforts on a Europe-wide scale are unlikely to reduce further spread of B. salamandrivorans and prevent it from eventually reaching centers of caudate endemism in southern Europe. That probability is highlighted by the previous money flow and resulting projects conducted in those areas (Italy, Portugal, Spain, and the entire Balkan Peninsula), which did not surpass €400,000 and 2 projects per country (Figure 1; Appendix 2 Table).
Any serious attempt to control an EID requires sufficient resources, in this case at least those necessary to support effective surveillance and mitigation actions as required by the AHL. Certainly, the risk B. salamandrivorans poses to Europe’s unique amphibian biodiversity and its potential of inducing a trophic cascade (42) should urge investment in resources to develop sustainable mitigation strategies (24,25). That risk extends beyond Europe to other yet naive regions such as northern Africa and North and South America. Resources invested in Europe automatically lead not only to previously mentioned actions but simultaneously follow approaches like One Health and reduce risk in and spillover to nonaffected countries (43). The time is now to enhance resources against the pathogen; once EIDs become widely established, enormous efforts are required to slow down epidemics in the wild (26). Our concern is that this might also be the case for B. salamandrivorans. If the current known distribution is representative of the actual distribution, we might still be in a position to enact effective mitigation strategies. The shown inadequacy of monetary resources and the coupled lack of B. salamandrivorans control measures in the EU become more evident by comparing them with resources spent on controlling other AHL-listed pathogens with established surveillance and mitigation structures. For example, African swine fever (ASF), an emerging disease of both wildlife and livestock, receives broad attention by decision-makers. In this case, because the pathogens differ in their epidemiology, host range, and economic effects, ASF is not used as a biologic comparator but as a policy benchmark to illustrate the differences in political priorities, funding allocation, and how control measures are implemented under the same legislative framework. This level of attention has led to massive investments in elimination and education measures with estimated funds at ≈€215 million for 2014–2023 (44,45). In detail, those EU funds, through the European Food Safety Authority for example, supported scientific projects regarding disease risk assessments. In addition, they include the cofinancing of national surveillance plans by the European Commission, which include active surveillance activities such as culling, carcass disposal, or physical barriers to prevent disease spread and passive surveillance in domestic pigs and wild boars; population management of wild boars; and biosecurity measures (https://www.efsa.europa.eu/en/topics/topic/african-swine-fever#latest). However, during that period the EU invested only 0.9% (€900,000) of the ASF funding (≈€215 million) in B. salamandrivorans mitigation, clearly showing that the EU-wide responses established for ASF were not feasible for B. salamandrivorans. Despite some initiated activities in some countries on different administrative levels, those efforts remain far less structured and consistent than those activated for ASF.
Examining how the United States has dealt with the threat of B. salamandrivorans, where the pathogen is not yet known to occur, the need for the financial and structural efforts we advocate for Europe become even more apparent. The United States has invested >$2 million in surveillance for B. salamandrivorans in wild amphibian populations (>100,000 samples) and has invested >$7 million in fundamental research, including estimating susceptibility of native species, investigating and modeling possible transmission dynamics in amphibian communities, evaluating disease management strategies, and using decision science to establish a response framework if emergence occurs (46–49). In comparison, ≈€1.2 million was invested in the EU in total (of which ≈66% included direct funding for research); 11,743 samples were tested in B. salamandrivorans–naive EU countries by May 2025 (Appendix 1 Table).
The North American Bsal Task Force serves as the unifying entity for that continent, bringing together scientists and stakeholders to identify research and policy needs and advocate for funding (49). The United States does not have specific legislation, such as the European Health Law, to direct funding toward B. salamandrivorans surveillance and response. Instead, funding has primarily come from state natural resource agencies, the US Geological Survey, and private conservation organizations that recognize the threat of this pathogen to native biodiversity. In addition, the US National Science Foundation has awarded funding for fundamental research on B. salamandrivorans and the possible threat of pathogen invasion through the captive amphibian trade. The availability of government funding for B. salamandrivorans research in the future, however, is uncertain, because many science-based programs are being cut by the current US administration. Recently, the threat of B. salamandrivorans and other pathogens has gained the attention of the US amphibian pet trade industry. US businesses overwhelmingly support the healthy trade of amphibians, and US consumers are willing to pay 75% more for pet amphibians that are not infected with B. salamandrivorans or other pathogens (50). This interest and demand led to the recent creation of the nonprofit Healthy Trade Institute, Inc., which is launching a healthy trade certification program for pet amphibians.
Thus, the threat of B. salamandrivorans is recognized by both public and private sectors in the United States, and proactively investing resources is agreed upon as essential to reducing likelihood of emergence and responding effectively if the pathogen is detected. Of note, research and policy have been closely linked in the United States; this strategy has proven highly effective in lessening the import by trade of known, and suspected, amphibian vectors of B. salamandrivorans (51).
In conclusion, we highlighted the financial gaps existing between livestock (ASF) and wildlife-only (B. salamandrivorans) diseases and uncovered funding differences between the United States, a country naive to B. salamandrivorans, and the EU, in which a few countries have reported B. salamandrivorans outbreaks. We call for the EU to take responsibility in amphibian conservation by implementing and enforcing the Animal Health Law across EU member states and their administrative units regarding B. salamandrivorans control measures in the wild and in captive facilities. Control measures in captive facilities especially include implementing improved biosecurity in the amphibian trade (i.e., following clean trade strategies) through mandatory health certification for amphibians, which remains largely absent in most member states despite its potential for effectively limiting pathogen introduction and spread.
These steps would require structural funding to (at the very least) cover costs of monitoring and activities inscribed in B. salamandrivorans action plans, to limit the effect of the wildlife-only EID on biodiversity in Europe in line with the European biodiversity strategy 2030 (52). In addition, it would involve fostering exchange between agencies funding fundamental research and those who are responsible for conservation action. Establishing clear and complementary funding connections would help to ensure that science informs action without reducing research to a mere service function. Subsequently, such coordination translates scientific findings into effective biodiversity management and disease control policies on all administrative scales, as well as in areas with high salamander diversity (especially southern Europe) and to species highly susceptible but naive to B. salamandrivorans, as already analyzed in the Batrachochytrium salamandrivorans Action Plan in Europe (24). To achieve this goal, we urgently call for a dialogue between policymakers, researchers, and stakeholders discussing and defining funding, coordination and implementation strategies, and priorities. For this urgent dialogue, we propose a coordinated EU-level program in the range of €6–10 million over 5–7 years focusing on 5 major components (Table) as a realistic starting point, based on preventive investment levels in comparable jurisdictions. (The United States has invested approximately $9 million in preventive B. salamandrivoranssurveillance and research despite the absence of confirmed cases.)
As we face the reality that the EU and its member states lack action against wildlife-only diseases such as chytridiomycosis, we urge more support in prevention measures such as campaigns to raise awareness, rapid-response systems, collaborative conservation actions, the implementation and maintenance of a B. salamandrivorans working group in Europe, and research in both hosts and pathogen following the efforts made in the United States to properly address the current disease emergency. Those actions are necessary and applicable to all known and yet unknown wildlife EIDs to elevate their recognition to that given to other wildlife diseases and to fulfill Europe’s obligation to One Health. In contrast, failing to address wildlife diseases risks undermining decades of conservation efforts and accelerating biodiversity loss across Europe. Because wildlife pathogens can spread rapidly across borders and ecosystems, coordinated prevention and surveillance are essential at the European level. Recognizing wildlife disease management as an integral component of One Health policy is critical to safeguarding biodiversity, ecosystem stability, and long-term environmental health.
Dr. Böning is a faculty member of the geobotany department at Trier University. His main interests include disease ecology and species conservation in amphibians and reptiles.
References
- Longshaw M. Diseases of crayfish: a review. J Invertebr Pathol. 2011;106:54–70. DOIPubMedGoogle Scholar
- Blehert DS. Fungal disease and the developing story of bat white-nose syndrome. PLoS Pathog. 2012;8:
e1002779 . DOIPubMedGoogle Scholar - Lorch JM, Knowles S, Lankton JS, Michell K, Edwards JL, Kapfer JM, et al. Snake fungal disease: an emerging threat to wild snakes. Philos Trans R Soc Lond B Biol Sci. 2016;371:
20150457 . DOIPubMedGoogle Scholar - Fisher MC, Garner TWJ. Chytrid fungi and global amphibian declines. Nat Rev Microbiol. 2020;18:332–43. DOIPubMedGoogle Scholar
- Daszak P, Cunningham AA, Hyatt AD. Emerging infectious diseases of wildlife—threats to biodiversity and human health. Science. 2000;287:443–9. DOIPubMedGoogle Scholar
- Springborn MR, Weill JA, Lips KR, Ibáñez R, Ghosh A. Amphibian collapses increased malaria incidence in Central America. Environ Res Lett. 2022;17:
104012 . DOIGoogle Scholar - Frank EG. The economic impacts of ecosystem disruptions: costs from substituting biological pest control. Science. 2024;385:
eadg0344 . DOIPubMedGoogle Scholar - Vaeßen S, Hollert H. Impacts of the North American signal crayfish (Pacifastacus leniusculus) on European ecosystems. Environ Sci Eur. 2015;27:33. DOIPubMedGoogle Scholar
- Jussila J, Edsman L, Maguire I, Diéguez-Uribeondo J, Theissinger K. Money kills native ecosystems: European crayfish as an example. Front Ecol Evol. 2021;9:
648495 . DOIGoogle Scholar - Weinländer M, Füreder L. Native and alien crayfish species: do their trophic roles differ? Freshw Sci. 2016;35:1340–53. DOIGoogle Scholar
- Hoyt JR, Kilpatrick AM, Langwig KE. Ecology and impacts of white-nose syndrome on bats. Nat Rev Microbiol. 2021;19:196–210. DOIPubMedGoogle Scholar
- Zipkin EF, DiRenzo GV, Ray JM, Rossman S, Lips KR. Tropical snake diversity collapses after widespread amphibian loss. Science. 2020;367:814–6. DOIPubMedGoogle Scholar
- Buttke DE, Decker DJ, Wild MA. The role of One Health in wildlife conservation: a challenge and opportunity. J Wildl Dis. 2015;51:1–8. DOIPubMedGoogle Scholar
- Adisasmito WB, Almuhairi S, Behravesh CB, Bilivogui P, Bukachi SA, Casas N, et al.; One Health High-Level Expert Panel (OHHLEP). One Health: a new definition for a sustainable and healthy future. PLoS Pathog. 2022;18:
e1010537 . DOIPubMedGoogle Scholar - World Organisation for Animal Health. OIE Wildlife Health Framework: protecting wildlife health to achieve One Health [cited 2023 Nov 15]. https://www.woah.org/fileadmin/Home/eng/Internationa_Standard_Setting/docs/pdf/WGWildlife/A_Wildlifehealth_conceptnote.pdf
- Ghai RR, Wallace RM, Kile JC, Shoemaker TR, Vieira AR, Negron ME, et al. A generalizable One Health framework for the control of zoonotic diseases. Sci Rep. 2022;12:8588. DOIPubMedGoogle Scholar
- Cunningham AA, Daszak P, Wood JLN. One Health, emerging infectious diseases and wildlife: two decades of progress? Philos Trans R Soc Lond B Biol Sci. 2017;372:
20160167 . DOIPubMedGoogle Scholar - Scheele BC, Pasmans F, Skerratt LF, Berger L, Martel A, Beukema W, et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science. 2019;363:1459–63. DOIPubMedGoogle Scholar
- Luedtke JA, Chanson J, Neam K, Hobin L, Maciel AO, Catenazzi A, et al. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature. 2023;622:308–14. DOIPubMedGoogle Scholar
- Martel A, Blooi M, Adriaensen C, Van Rooij P, Beukema W, Fisher MC, et al. Wildlife disease. Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science. 2014;346:630–1. DOIPubMedGoogle Scholar
- Gray MJ, Carter ED, Piovia-Scott J, Cusaac JPW, Peterson AC, Whetstone RD, et al. Broad host susceptibility of North American amphibian species to Batrachochytrium salamandrivorans suggests high invasion potential and biodiversity risk. Nat Commun. 2023;14:3270. DOIPubMedGoogle Scholar
- DiRenzo GV, Longo AV, Muletz-Wolz CR, Pessier AP, Goodheart JA, Lips KR. Plethodontid salamanders show variable disease dynamics in response to Batrachochytrium salamandrivorans chytridiomycosis. Biol Invasions. 2021;23:2797–815. DOIGoogle Scholar
- Martel A, Spitzen-van der Sluijs A, Blooi M, Bert W, Ducatelle R, Fisher MC, et al. Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proc Natl Acad Sci U S A. 2013;110:15325–9. DOIPubMedGoogle Scholar
- Gilbert MJ, Spitzen-van der Sluijs A, Canessa S, Bosch J, Cunningham AA, Grasselli E, et al. Mitigating Batrachochytrium salamandrivorans in Europe. Batrachochytrium salamandrivorans Action Plan for European urodeles [cited 2023 Nov 15]. http://bsaleurope.com/wp-content/uploads/2021/03/Bsal-Action-Plan.pdf
- Thomas V, Wang Y, Rooij PV, Verbrugghe E, Baláž V, Bosch J, et al. Mitigating Batrachochytrium salamandrivorans in Europe. Amphib-Reptil. 2019;40:265–90. DOIGoogle Scholar
- Martel A, Vila-Escale M, Fernández-Giberteau D, Martinez-Silvestre A, Canessa S, Van Praet S, et al. Integral chain management of wildlife diseases. Conserv Lett. 2020;13:
e12707 . DOIGoogle Scholar - Akçakaya HR, Neam K, Hobin L, Lötters S, Martel A, Pasmans F. Assessing the extinction risks of amphibians impacted by infectious diseases. Biol Conserv. 2023;284:
110205 . DOIGoogle Scholar - Böning P, Plewnia A, Virgo J, Adam J, Banowski N, Bleidißel S, et al. Salamander plague: characterization and current situation in Germany. Recommendation for action [in German]. Z Feldherpetol. 2024;31:1–38.
- Sabino-Pinto J, Veith M, Vences M, Steinfartz S. Asymptomatic infection of the fungal pathogen Batrachochytrium salamandrivorans in captivity. Sci Rep. 2018;8:11767. DOIPubMedGoogle Scholar
- Nguyen TT, Van Nguyen T, Ziegler T, Pasmans F, Martel A. Trade in wild anurans vectors the urodelan pathogen Batrachochytrium salamandrivorans into Europe. Amphib-Reptil. 2018;38:554–6. DOIGoogle Scholar
- European Union. Regulation (EU) 2016/429 of the European Parliament and of the Council of 9 March 2016 on transmissible animal diseases and amending and repealing certain acts in the area of animal health (‘Animal Health Law’) [cited 2023 Nov 11]. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32016R0429
- Sun D, Ellepola G, Herath J, Meegaskumbura M. The two chytrid pathogens of amphibians in Eurasia-climatic niches and future expansion. BMC Ecol Evol. 2023;23:26. DOIPubMedGoogle Scholar
- Beukema W, Erens J, Schulz V, Stegen G, Spitzen-van der Sluijs A, Stark T, et al. Landscape epidemiology of Batrachochytrium salamandrivorans: reconciling data limitations and conservation urgency. Ecol Appl. 2021;31:
e02342 . DOIPubMedGoogle Scholar - Lastra González D, Baláž V, Solský M, Thumsová B, Kolenda K, Najbar A, et al. Recent findings of potentially lethal salamander fungus Batrachochytrium salamandrivorans. Emerg Infect Dis. 2019;25:1416–8. DOIPubMedGoogle Scholar
- Baláž V, Solský M, Lastra González D, Havlíková B, Zamorano JG, González C, et al. First survey of the pathogenic fungus Batrachochytrium salamandrivorans in wild and captive amphibians in the Czech Republic. Salamandra (Frankf). 2018;54:87–91.
- Fitzpatrick LD, Pasmans F, Martel A, Cunningham AA. Epidemiological tracing of Batrachochytrium salamandrivorans identifies widespread infection and associated mortalities in private amphibian collections. Sci Rep. 2018;8:13845. DOIPubMedGoogle Scholar
- Grasselli E, Bianchi G, Dondero L, Marchianò V, Carafa M, Perrone M, et al. First screening for Batrachochytrium salamandrivorans (Bsal) in wild and captive salamanders from Italy. Salamandra (Frankf). 2019;55:124–6.
- Marquis O, Miaud C, Gibault C, Chai N. A first screening of chytrid fungus Batrachochytrium in amphibians in French zoos. Int Zoo Yearb. 2019;53:217–26. DOIGoogle Scholar
- Kostanjšek R, Turk M, Vek M, Guitérrez-Aguirre I, Cimerman NG. First screening for Batrachochytrium dendrobatidis, B. salamandrivorans and ranavirus infections in wild and captive amphibians in Slovenia. Salamandra (Frankf). 2021;57:162–6.
- Thumsová B, Bosch J, Martínez-Silvestre A. Incidence of emerging pathogens in the legal and illegal amphibian trade in Spain. Herpetol Notes. 2021;14:777–84.
- Kelly M, Pasmans F, Muñoz JF, Shea TP, Carranza S, Cuomo CA, et al. Diversity, multifaceted evolution, and facultative saprotrophism in the European Batrachochytrium salamandrivorans epidemic. Nat Commun. 2021;12:6688. DOIPubMedGoogle Scholar
- Laking AE, Li Z, Goossens E, Miñarro M, Beukema W, Lens L, et al. Salamander loss alters litter decomposition dynamics. Sci Total Environ. 2021;776:
145994 . DOIPubMedGoogle Scholar - Plowright RK, Ahmed AN, Coulson T, Crowther TW, Ejotre I, Faust CL, et al. Ecological countermeasures to prevent pathogen spillover and subsequent pandemics. Nat Commun. 2024;15:2577. DOIPubMedGoogle Scholar
- Palencia P, Blome S, Brook RK, Ferroglio E, Jo YS, Linden A, et al. Tools and opportunities for African swine fever control in wild boar and feral pigs: a review. Eur J Wildl Res. 2023;69:69. DOIGoogle Scholar
- European Union. Animal health: the EU’s fight against African swine fever [cited 2023 Nov 15]. https://food.ec.europa.eu/document/download/562e4071-2415-40c7-af00-c848ee1ab7bc_en?filename=ad_control-measures_asf_factsheet_2023.pdf
- Gray MJ, Lewis JP, Nanjappa P, Klocke B, Pasmans F, Martel A, et al. Batrachochytrium salamandrivorans: the North American response and a call for action. PLoS Pathog. 2015;11:
e1005251 . DOIPubMedGoogle Scholar - Waddle JH, Grear DA, Mosher BA, Grant EHC, Adams MJ, Backlin AR, et al. Batrachochytrium salamandrivorans (Bsal) not detected in an intensive survey of wild North American amphibians. Sci Rep. 2020;10:13012. DOIPubMedGoogle Scholar
- Gray MJ, Carter ED, Piovia-Scott J, Cusaac JPW, Peterson AC, Whetstone RD, et al. Broad host susceptibility of North American amphibian species to Batrachochytrium salamandrivorans suggests high invasion potential and biodiversity risk. Nat Commun. 2023;14:3270. DOIPubMedGoogle Scholar
- Olson DH, Grant EHC, Bletz M, Piovia-Scott J, Lesbarrères D, Kerby JL, et al. Preparing for a Bsal invasion into North America has improved multi-sector readiness. Front Amphib Reptile Sci. 2024;2:
1347541 . DOIGoogle Scholar - Cavasos K, Adhikari RK, Poudyal NC, Brunner JL, Warwick AR, Gray MJ. Understanding the demand for and value of pathogen‐free amphibians to US pet owners. Conserv Sci Pract. 2023;5:
e12995 . DOIGoogle Scholar - Conelly PJ, Ross N, Stringham OC, Eskew EA. United States amphibian imports pose a disease risk to salamanders despite Lacey act regulations. Commun Earth Environ. 2023;4:351. DOIGoogle Scholar
- European Union. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions: EU Biodiversity Strategy for 2030—bringing nature back into our lives [cited 2026 Jun 19]. https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex:52020DC0380
- European Union. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora [cited 2026 Jun 25]. http://data.europa.eu/eli/dir/1992/43/2013-07-01
Figures
Table
Cite This ArticleOriginal Publication Date: June 25, 2026
Table of Contents – Volume 32, Number 7—July 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Philipp Böning, Department of Geobotany, Trier University, Universitätsring 15, 54296 Trier, Germany
Top