Volume 32, Number 7—July 2026
Dispatch
Angiostrongylus cantonensis Rat Lungworm Detected in Rats, Madagascar, 2022–2023
Cite This Article
Citation for Media
Abstract
Angiostrongylus cantonensis, the rat lungworm, is a zoonotic parasite that causes eosinophilic meningitis in humans; the parasite is maintained in rat definitive hosts and transmitted through gastropod intermediate hosts. We report A. cantonensis prevalence and mitochondrial genome from oral swab specimens from rats in Madagascar, supporting swabs for noninvasive detection of this parasite.
Angiostrongylus cantonensis, the rat lungworm, is a zoonotic nematode and a leading cause of eosinophilic meningitis in humans and other mammals (1). Originally described in rats in China in 1935, A. cantonensis lungworm is now reported in >30 countries globally (2). Distribution has expanded because of globalization, human activity, and range expansion of definitive and intermediate hosts (2,3).
Adult A. cantonensis worms inhabit the pulmonary arteries of rats (definitive hosts), including black rats (Rattus rattus) and brown rats (Rattus norvegicus), where eggs hatch into first-stage larvae shed in feces. Those larvae infect intermediate hosts such as snails and slugs, where they develop into third-stage larvae and can be transmitted via paratenic hosts (including reptiles and amphibians), where larvae can persist for extended periods, and through transient hosts (e.g., crustaceans and insects), where larvae survive for only a short time (4). Rats acquire infection by ingesting those hosts, completing the life cycle (5). Humans are infected accidentally through consumption of raw or undercooked intermediate or paratenic hosts or via contaminated food and water (6). After ingestion, the larvae migrate from the intestines to the brain, causing meningitis and neurologic complications (4,7).
Madagascar is one of the world’s most expansive biodiversity hotspots, where humans and an exceptional diversity of endemic wildlife frequently interact with nonnative animals across the island (8). Although A. cantonensis worms have been reported in the region (9), genomic data and epidemiologic insights remain limited. In this study, we used metagenomic sequencing of oral swabs to characterize mitochondrial genomes and estimate prevalence of A. cantonensis lungworm in black rats from southeastern Madagascar.
Figure 1
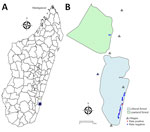
Figure 1. Geographic location of Manombo Special Reserve and spatial distribution of sampled rats in a study of Angiostrongylus cantonensisrat lungworm detection in rats in southeastern Madagascar, 2022–2023. A) Location...
As part of a broader One Health study (10), we collected oral swab samples from 125 wild black rats that were captured, anesthetized, and euthanized during October 2022 and July 2023 in the Manombo Special Reserve (MSR), southeastern Madagascar (Figure 1, panels A, B). The samples were collected in 3 different habitats: littoral forest (n = 76 samples), lowland rainforest (n = 22 samples), and village-edge (n = 27 samples).
We conducted DNA extraction by using the Roche HighPure Viral Nucleic Acid Kit (Roche, https://www.roche.com). We followed our previously published pipeline for library preparation, sequencing, bioinformatic processing, quality trimming, de novo assembly, and contig screening by using standard tools and the National Center for Biotechnology Information (NCBI) mitochondrial RefSeq database (11). We annotated mitochondrial genomes by using reference genomes of A. cantonensis (NCBI Sequence Read Archive accession no. GQ398121) with manual curation.
For phylogenetic analysis, we aligned complete mitochondrial genomes with publicly available A. cantonensis sequences. We constructed a maximum-likelihood tree by using IQ-Tree 2 (12) to determine the genetic relationships of our samples and haplotype placement.
Because metagenome-assembled A. cantonensis mitogenomes were highly similar, we mapped raw reads to a representative mitogenome by using CoverM (13) with a minimum read identity of 95%. We used a read coverage threshold of >75% to determine presence or absence of A. cantonensis genomes in each sample, providing a high-confidence proxy for infection status and calculated prevalence accordingly. To identify predictors of infection, we performed a generalized linear model in R version 4.4.1 (The R Project for Statistical Computing, https://www.r-project.org), testing age, sex, location, body condition, weight, and year of sampling.
All procedures were approved by Duke University Institutional Animal Care and Use committee (approval no. A075‐23‐03), Zoo New England ethics board, and the Madagascar Ministry of the Environment (approval nos. 286/22 and 215/23/MEDD/SG/DGGE/DAPRNE/SCBE.Re). We deposited all raw sequencing data in the NCBI Sequence Read Archive. Taxonomic classification revealed that 26.26%–51.43% of reads per sample were assigned to known taxa, including bacteria (15.01%–47.72%), eukaryotes (0.53%–4.62%), and viruses (0.04%–18.78%). Viral sequences mainly comprised anelloviruses and bacteriophages (10).
Figure 2

Figure 2. Mitochondrial genome organization and sequence coverage maps of Angiostrongylus cantonensis rat lungworm from black rat oral swab specimens in southeastern Madagascar, 2022–2023.
Figure 3

Figure 3. Whole-mitogenome maximum likelihood phylogenetic tree of complete sequences available for Angiostrongylus cantonensis rat lungworm and 3 related Angiostrongylus species along with the sequences identified in study of ...
We detected 5 complete A. cantonensis mitochondrial genomes from black rat oral swab specimens, de novo assembled with high coverage (GenBank accession nos. PX571103–7) (Figure 2). All 5 mitochondrial genomes were 13,503 nt in length with 26.8% guanine and cytosine content, in comparison with published A. cantonensis mitogenomes that were 13,497–13,519 nt in length with 26.7%–26.8% guanine and cytosine content. The mitogenomes from Madagascar revealed 99.96%–100% sequence similarity among themselves and 96.44%–99.99% similarity compared with global sequences. Of note, the genomes from Madagascar demonstrated >99.98% similarity to sequences from Valencia, Spain (GenBank accession no. PP748572). Phylogenetic analysis placed all sequences from Madagascar within clade II, grouping with the Val-II haplotype and clustering with the haplotype Ac8 from Brazil (Figure 3) (14).
A. cantonensis prevalence in black rat oral swabs was determined to be 10.4% (95% CI 6.2%–16.9%; 13/125 samples). All positive rats were from the littoral forest of the MSR (Figure 1, panel B). Positive cases were detected in both sampling years, with no significant difference in prevalence between years: 9.67% (95% CI 4.5%–19.6%; 6/62) in 2022 and 11.11% (95% CI 5.5%–21.4%; 7/63) in 2023. Logistic regression analysis identified age as the only significant predictor of infection. Young rats had significantly lower odds of infection compared to adults (odds ratio 0.13, 95% CI 0.02–0.77; p = 0.03) (Table). We observed no significant associations for sex, body condition, weight, or year of sampling (Table).
The 5 mitogenomes we obtained were nearly identical to each other (99.96%–100% similarity), indicating a highly conserved parasite population within the MSR littoral forest and low genetic diversity of A. cantonensis lungworm in Madagascar, which is similar to another report (3). Phylogenetic analysis further demonstrated that all the sequences cluster within clade II and specifically group with the Val-II haplotype, previously reported in Europe and associated with haplogroup Ac8 from Brazil (14). That pattern supports the hypothesis of recent or historical introductions mediated by human activities, particularly through the movement of invasive rats and intermediate hosts via trade and transport networks (3,14).
The overall prevalence in rats in the current study was 10.4% (95% CI 6.2%–16.9%; 13/125 rats), higher than the 2.7% prevalence previously reported in rats from central-eastern Madagascar (n = 78) (9). All positive rats were from the littoral forests. Host-related factors also contributed to infection patterns. Age was the only significant predictor; young rats showed markedly lower odds of infection compared with adults. That finding reflects cumulative exposure over time, because older rats have more opportunities to encounter infected intermediate or paratenic hosts (15). We observed no significant effects for sex, body condition, weight, or year, suggesting that exposure-driven processes might be more necessary than intrinsic host factors in shaping infection risk. No positive detections were observed in the lowland rainforest or village-edge habitats, potentially because of intermediate host distribution (5).
This study provides metagenomic-based mitochondrial genome characterization and prevalence estimates of A. cantonensis lungworm from black rats in southeastern Madagascar. Because of the parasite’s life cycle, oral swab specimens likely capture first-stage larvae expelled from the respiratory tract, which are known to be present in the oral cavity during patent infections. However, because necropsy data were not systematically collected for parasitologic examination, we could not assess the relationship between larval detection in oral swab specimens and adult worm burden in the pulmonary vasculature. Future studies combining oral swab specimens with necropsy or quantitative parasitologic methods would help validate this relationship. Nevertheless, our findings demonstrate that complete A. cantonensis mitochondrial genomes can be recovered from oral swab specimens, supporting their use as a complementary, minimally invasive approach for molecular detection and genomic characterization of this parasite.
Dr. Randrianarisoa is a wildlife veterinarian and laboratory manager at Mahaliana Labs in Antananarivo, Madagascar. His research focuses on zoonotic pathogens, antimicrobial resistance, and wildlife health at the human–animal–environment interface in Madagascar.
Acknowledgments
We gratefully acknowledge Zoo New England for their continued funding support and dedication to One Health and biodiversity conservation research at MSR.
We also extend our appreciation to the local guides, cooks, and Madagascar National Parks staff who assisted with fieldwork during the 2022 and 2023 sampling efforts. We thank Health In Harmony-Fahasalamana Mirindra for fostering strong relationships with communities, providing communication about our research activities, and serving as an essential bridge between researchers and community members. Our sincere gratitude goes to each village’s Mpanjaka (the King or Queen of the village) and the communities of all 31 villages for granting us access to their forests. We likewise acknowledge the Centre ValBio research technicians who contributed to rodent sampling in both years. We appreciate the support of the Madagascar Ministry of the Environment and Sustainable Development and the Duke University Institutional Animal Care and Use Committee for granting research permissions. Finally, we thank Madagascar Institute for the Conservation of Tropical Ecosystems, with special acknowledgment to Tiana Vololona, for their valuable assistance in coordinating research logistics and permits.
The sequences described in this study are deposited in GenBank (accession nos. PX571103–7). The raw reads for the black rat oral swab samples are deposited in the NCBI Sequence Read Archive (BioProject no. PRJNA1290322; Biosample nos. SAMN49916503, SAMN49916584, SAMN49916600, SAMN49916604, and SAMN49916608; Sequence Read Archive accession nos. SRR34496630, SRR34496775, SRR34496757, SRR34496753, and SRR34496748).
This study was supported in part by a grant from the Wild Animal Health Fund of the American Association of Zoo Veterinarians to Zoo New England and Mahaliana Labs. Additional funding for E.N.P. was provided by Duke University.
References
- Centers for Disease Control and Prevention. Angiostrongyliasis cantonensis. 2019 [cited 2025 Nov 10]. https://www.cdc.gov/dpdx/angiostrongyliasis_can/index.html
- Barratt J, Chan D, Sandaradura I, Malik R, Spielman D, Lee R, et al. Angiostrongylus cantonensis: a review of its distribution, molecular biology and clinical significance as a human pathogen. Parasitology. 2016;143:1087–118. DOIPubMedGoogle Scholar
- Červená B, Modrý D, Fecková B, Hrazdilová K, Foronda P, Alonso AM, et al. Low diversity of Angiostrongylus cantonensis complete mitochondrial DNA sequences from Australia, Hawaii, French Polynesia and the Canary Islands revealed using whole genome next-generation sequencing. Parasit Vectors. 2019;12:241. DOIPubMedGoogle Scholar
- Galán-Puchades MT, Gómez-Samblás M, Osuna A, Sáez-Durán S, Bueno-Marí R, Fuentes MV. Update on the first finding of the rat lungworm, Angiostrongylus cantonensis, in Rattus spp. in continental Europe, Valencia, Spain, 2022. Pathogens. 2023;12:567. DOIPubMedGoogle Scholar
- Cowie RH. Biology, systematics, life cycle, and distribution of Angiostrongylus cantonensis, the cause of rat lungworm disease. Hawaii J Med Public Health. 2013;72(Suppl 2):6–9.PubMedGoogle Scholar
- Wang QP, Lai DH, Zhu XQ, Chen XG, Lun ZR. Human angiostrongyliasis. Lancet Infect Dis. 2008;8:621–30. DOIPubMedGoogle Scholar
- Li T mei, Liu Y hua, Fang W, Zhao S hua, Li T, Jiang L, et al. Monitoring the trends of Angiostrongylus cantonensis infection in humans and Pomacea spp. snails in Dali, Yunnan, China, 2007–2021. PLOS Neglected Tropical Diseases. 2025;19:e0013065.
- Rasambainarivo F, Goodman SM. Disease risk to endemic animals from introduced species on Madagascar. Fowler’s Zoo and Wild Animal Medicine Current Therapy. 2019;9:292–7. DOIGoogle Scholar
- Maminirina LA, Bodoarison ZI, Rajerison M, Ferdinand S, Ramasindrazana B. Angiostrongylus cantonensis lungworms in definitive and intermediate hosts, Madagascar, 2024. Emerg Infect Dis. 2025;31:2054–6. DOIPubMedGoogle Scholar
- Paietta EN, Johnston RA, Randrianarisoa SF, DeSisto CMM, Kraberger S, Martin D, et al. Host-anellovirus interactions in an island ecosystem: non-human primates and rodents in Madagascar harbour diverse, rich anellovirus populations. Microb Genom. 2026;12:
001681 . DOIPubMedGoogle Scholar - Paietta EN, Randrianarisoa SF, Razanamahenina TT, Ramboninarimalala A, Raherinirina TG, Raveloson L, et al. Complete mitochondrial DNA sequences of Eliurus webbi and Eliurus minor, endemic tufted-tailed rats in Madagascar. Mitochondrial DNA B Resour. 2026;11:561–6. DOIPubMedGoogle Scholar
- Minh BQ, Schmidt HA, Chernomor O, Schrempf D, Woodhams MD, von Haeseler A, et al. IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol Biol Evol. 2020;37:1530–4. DOIPubMedGoogle Scholar
- Aroney STN, Newell RJP, Nissen JN, Camargo AP, Tyson GW, Woodcroft BJ. CoverM: read alignment statistics for metagenomics. Bioinformatics. 2025;41:
btaf147 . DOIPubMedGoogle Scholar - Gómez-Samblás M, Navarro-Dominguez B, Sáez-Durán S, Osuna A, Bueno-Marí R, Galán-Puchades MT, et al. Analysis of the mitochondrial genome to determine the origins and pathways of entry of Angiostrongylus cantonensis in continental Europe (Valencia, Spain). Parasitology. 2024;151:1364–70. DOIPubMedGoogle Scholar
- Rivory P, Bedoya-Pérez M, Ward MP, Šlapeta J. Older urban rats are infected with the zoonotic nematode Angiostrongylus cantonensis. Curr Res Parasitol Vector Borne Dis. 2024;5:
100179 . DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: June 23, 2026
Table of Contents – Volume 32, Number 7—July 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Santatriniaina F. Randrianarisoa, Mahaliana Labs, Lot II B 55 G Amboditsiry, Antananarivo 101, Madagascar
Top