Volume 6, Number 6—December 2000
Research
Risk Factors for Otitis Media and Carriage of Multiple Strains of Haemophilus influenzae and Streptococcus pneumoniae
Cite This Article
Citation for Media
Abstract
We studied genetic diversity in Streptococcus pneumoniae and Haemophilus influenzae in throat culture isolates from 38 children attending two day-care centers in Michigan. Culture specimens were collected weekly; 184 S. pneumoniae and 418 H. influenzae were isolated from the cultures. Pulsed-field gel electrophoresis identified 29 patterns among the S. pneumoniae isolates and 87 among the H. influenzae isolates. Of the cultures, 5% contained multiple genetic types of S. pneumoniae, and 43% contained multiple types of H. influenzae. Carriage of multiple H. influenzae isolates, which was associated with exposure to smoking, history of allergies, and age 36 to 47 months, may increase risk for otitis media in children.
Acute otitis media is the most frequently diagnosed bacterial illness in young children in the United States, resulting in an annual economic cost of approximately $3.8 billion per year (1). Streptococcus pneumoniae and nontypable Haemophilus influenzae are the bacterial species most frequently associated with otitis media in young children, and nasopharyngeal colonization with these bacteria is associated with its development (2-6).
Simultaneous carriage of more than one genetically distinct isolate of H. influenzae has been observed in Australian Aboriginal children (7) and in children attending day-care centers (8), two groups at increased risk for otitis media (4,7,9-12). Carriage of multiple genetic types of H. influenzae may result from inadequate clearance of colonizing bacteria or from exposure to multiple circulating strains (4,7). Concurrent colonization with more than one serotype of S. pneumoniae has also been reported (13), but the degree of colonization with multiple genetic types of S. pneumoniae has not been investigated. In addition, the association between risk factors for otitis media and colonization with multiple types of H. influenzae and S. pneumoniae isolates has not been examined.
We examined genetic diversity among S. pneumoniae and H. influenzae isolates colonizing 38 children attending two Michigan day-care centers. We also investigated the degree to which the bacteria were shared among children attending these centers and the relationship between risk factors for otitis media and carriage of multiple isolates.
Study Population
Thirty-eight children from two Michigan day-care centers participated in the study. One hundred sixty children attended day-care center 1 full time, divided into nine classrooms by age and developmental level. At least one child from each classroom participated in the study, with the exception of the classroom for children >5 years of age. Fifty-five children attended day-care center 2 full time, divided into six classrooms. A total of 12% of the children from day-care center 1 and 33% from day-care center 2 participated in the study; at least one child from each classroom participated.
Study Protocol
Participants were recruited from day-care center 1 in February 1998 and from day-care center 2 in June 1998. Parents signed consent forms allowing four throat cultures to be collected from the children over a 3-week period. Parents were also asked to complete a short questionnaire assessing history of and risk factors for otitis media, antibiotic use, and demographic characteristics. The study was approved by the University of Michigan Health Sciences Institutional Review Board.
Throat cultures were collected once a week for 3 weeks from each child, for a maximum of four cultures per child. At the time of each culture, the child's teacher was asked if the child had been ill in the preceding week, and if so, with what symptoms. The parents of children who had been ill were asked to describe the child's symptoms and whether an antibiotic was given.
Bacteriologic Methods
Throat swabs were collected with rayon-tipped swabs (VWR Scientific, Westchester, PA), were placed in 500 µL Brain Heart Infusion (BHI) medium, and were transported on ice. Swabs were vortexed vigorously in the medium and then discarded. Five hundred µL sterile skim milk was added to each sample, and cultures were frozen at -80°C.
Samples were thawed at room temperature, vortexed briefly, and 100 µL of sample was streaked on each of two chocolate agar plates supplemented with 300 mg bacitracin (BBL, VWR Scientific) for H. influenzae selection and two 5% sheep blood agar plates supplemented with 2.5 µg gentamicin (BBL) for S. pneumoniae. Plates were incubated at 37°C with 5% CO2 for 24 hours to 48 hours. Five colonies were selected from each plate for a maximum of 10 colonies per bacterial species; colonies with differing morphologic features were selected. H. influenzae isolates were characterized by X- and V-factor dependence (BBL): 34% of the picked colonies were H. influenzae, 49% were H. parainfluenzae, and 17% were other species. Only H. influenzae isolates were characterized further. S. pneumoniae isolates were identified by optochin disc sensitivity (BBL).
DNA Isolation from H. influenzae and S. pneumoniae
H. influenzae and S. pneumoniae isolates were streaked from frozen stocks on chocolate agar plates and blood agar plates, respectively, and grown overnight at 37°C with 5% CO2. Bacterial growth was diluted to an optical density of 1.0 (610 nm; Turner, Barnstead/Thermodyne, Dubuque, IA) SP 830 spectrophotometer) in 3 mL PETT IV buffer (10 mM Tris-Cl, pH 7.6, 1 M NaCl). The bacterial suspension (350 µL for H. influenzae or 1 mL for S. pneumoniae) was centrifuged at 13,000 rpm for 30 seconds. Bacterial pellets were resuspended in 100 mL PETT IV buffer, 100 µL 0.8% InCert agarose (FMC Bioproducts, Rockland, ME) was added to each, and 20 mL plugs were pipetted on a weigh boat. Plugs were cooled at 4°C for 30 minutes and then incubated with 1 mL EC lysis buffer (6 mM Tris-Cl, pH 7.6; 1 M NaCl; 100mM EDTA, pH 8.0; 0.5% Brij-58; 0.2% deoxycholate; 0.5% N-lauroylsarcosine; 1 mg/mL lysozyme; and 20 µg/mL RNAse) for 4 hours at 37°C (H. influenzae) or 50°C (S. pneumoniae). The buffer was removed, and plugs were incubated with 200 µL ESP buffer (0.5 M EDTA, pH 8.0; 1% N-lauroylsarcosine; and 1 mg/mL Proteinase K) for 12 hours to 24 hours at 50°C. ESP buffer was removed, and plugs were incubated with 1 mL TE-phenylmethylsulphonyl fluoride (1 mM) for 2 hrs at 37°C. Plugs were then washed twice with 1 mL TE buffer and stored at 4°C in 1 mL fresh TE buffer.
DNA Digestion and Pulsed-Field Gel Electrophoresis
Single plugs were incubated 30 min in 100 µL 1X enzyme reaction buffer (Gibco-BRL, Rockville, MD) at 37°C. Buffer was removed, and plugs were incubated 4 to 6 hrs with 69 µL 1X reaction buffer plus 1 µL SmaI (Gibco-BRL). Following digestion, restriction buffer was removed, and plugs were incubated 15 min in 0.5X TBE buffer at room temperature. Plugs were melted briefly at 80°C and loaded on 1% SeaKem HGT agarose gels (FMC Bioproducts). DNA electrophoresis was performed for 16 hrs with 4- to 16-second switch times at 120 volts and 14°C, with the Bio-Rad CHEF-DR III System (Hercules, CA). Three of the H. influenzae isolates did not digest well with SmaI and were excluded from the analysis. Twenty plugs from each species were also digested with ApaI. Bacterial isolates whose restriction patterns appeared identical or nonidentical with SmaI digestion also appeared to be identical or nonidentical, respectively, with ApaI digestion.
Figure 1
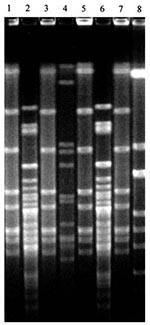
Figure 1. Four distinct Haemophilus influenzae isolates from one child during a sampling period. Lanes 1-8: H. influenzae isolates collected from child no. 32 at the third sampling period.
Figure 2
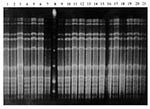
Figure 2. Example of S. pneumoniae isolates collected from child no. 37 during four sampling periods. Lane 1: Isolate collected at week 1; lanes 2-3: isolates collected at week 2; lanes 4-7, 9-13:...
Gels were incubated 1 to 4 hours with 20 µg/µL ethidium bromide, visualized under ultraviolet light, and photographed. The banding patterns were identified as either identical or different by the criteria of Tenover et al. (14). Identical isolates differed by no more than one band; closely related isolates by two or three bands; possibly related isolates by four to six bands; and different isolates by seven or more bands. All isolates of the same species collected from the same child were run on the same gel, so genetic fingerprints could be compared for all 3 weeks. For example, all H. influenzae isolates from child no. 32 were run on a single gel (Figure 1), while all S. pneumoniae isolates from child no. 37 were run on a single gel (Figure 2).
Analysis of Bacterial Sharing
Single isolates representing all observed pulsed-field gel electrophoresis (PFGE) patterns on the gels from individual children were run on a second gel to determine if particular patterns were shared among children in the day-care centers. Since not all unique H. influenzae patterns could be run on a single gel, individual H. influenzae gels were scanned into Adobe Photoshop (15) to permit comparison across separate gels. Any isolates that appeared identical in Adobe Photoshop were run together on a third gel for confirmation.
All statistics were performed with SAS (16). Fisher's exact test (two-tailed) was used to determine significant differences in the number of genetic types in day-care center 1 compared with day-care center 2. Univariate generalized estimating equation (GEE) logistic regression models were created to determine the risk factors for otitis media associated with carriage of multiple genetic types of H. influenzae or S. pneumoniae. The GEE SAS macro was used to control for correlation between multiple throat samples from the same child. Variables strongly associated with outcomes in univariate models were included in multivariate GEE models.
Colonization with S. pneumoniae and H. influenzae
We obtained 126 throat cultures from 38 children at two day-care centers. One hundred two (81%) of the cultures were positive for at least one H. influenzae isolate and 60 (48%) for at least one S. pneumoniae isolate. We obtained 418 H. influenzae isolates (average of 4.1/positive culture) and 184 S. pneumoniae isolates (average of 3.0/positive culture) from the 126 throat cultures.
PFGE analysis identified 87 patterns among the H. influenzae isolates and 29 patterns among the S. pneumoniae isolates. Two of the H. influenzae isolates appeared to be closely related, as their PFGE patterns differed by only two bands. Four other pairs of H. influenzae isolates were possibly related, as their banding patterns differed by four to six bands. All other H. influenzae isolates differed by seven or more bands. All nonidentical S. pneumoniae isolates differed by seven or more bands.
Fifty-six distinct H. influenzae patterns were observed in day-care center 1. Eight (14%) of those patterns were observed in more than one child during the study period. Thirty-one distinct H. influenzae patterns were observed in day-care center 2. Four (13%) patterns were observed in more than one child (Table 1). Thirteen distinct S. pneumoniae genetic patterns were observed in day-care center 1. Four (31%) patterns were observed in more than one child during the 3-week study period. Sixteen genetic patterns were observed in day-care center 2. Three (19%) of those patterns were observed in more than one child (Table 2).
Sharing of specific isolates varied from week to week and room to room within the day-care centers. Most shared isolates (68%) appeared in no more than two children during the study period. For example, in day-care center 1, the H. influenzae fingerprint pattern #7 appeared in children from five classrooms on three sampling occasions (Table 1).
Genetic Diversity of Isolates Carried by Individual Children
Fifty-four (43%) throat cultures contained two or more genetically distinct H. influenzae isolates, and children in day-care center 1 carried more genetically distinct isolates at a single time than children in day-care center 2 (p =.028). The children carried an average of 1.4 genetically distinct H. influenzae isolates per throat culture (range 0-5; Table 1) and 2.9 (range 0-8) genetically distinct isolates during the study period.
Six (5%) throat cultures contained two genetically distinct S. pneumoniae isolates, and none contained more than two (Table 2). On average, children carried fewer genetically distinct S. pneumoniae isolates per culture than the average number of H. influenzae isolates per culture (0.5 S. pneumoniae isolates vs. 1.4 H. influenzae isolates per culture); this difference was statistically significant (p <.001, t test). One child carried three genetically distinct S. pneumoniae isolates during the study period; no other child carried more than two isolates during the study period. Five of the children who carried more than one genetically distinct S. pneumoniae isolate simultaneously carried more than one genetically distinct H. influenzae isolate. Overall, 31 children (82%) carried multiple isolates at some point during the study.
Association with Risk Factors for Otitis Media
Children who were 36 to 47 months old, had allergies, or were exposed to smoking were more than twice as likely to carry two or more genetically distinct H. influenzae isolates at a time (Table 3). Children who had a history of frequent otitis media episodes were half as likely to carry two or more genetically distinct H. influenzae isolates. Children who used a pacifier were one-fifth as likely to carry two or more genetically distinct H. influenzae isolates as children who did not use a pacifier (Table 3). No exposure variables were associated with carriage of multiple distinct S. pneumoniae isolates.
In a multivariate GEE logistic regression model, exposure to smoking, having allergies, and being 36 to 47 months old were significantly associated with at least a twofold increase in carriage of two or more genetically distinct H. influenzae at a time (Table 4). Using a pacifier and having a history of frequent otitis media episodes were highly correlated with antibiotic use; therefore, only antibiotic use was included in the multivariate model. Even after the data were adjusted, children who had taken an antibiotic during the study were almost four times less likely to carry two or more genetically distinct H. influenzae isolates than children who had not taken an antibiotic (Table 4).
Multiple H. influenzae isolates are frequently present in the upper respiratory tracts of adults with cystic fibrosis (17) and chronic obstructive pulmonary disease (18). Multiple genetic types were identified in 44% of the throat cultures in our study, a rate similar to that observed in cultures obtained from three Australian Aboriginal children (7), but considerably higher than that recorded in previous studies (8,19,20).
The high genetic diversity we observed in individual children may be associated with use of throat culture specimens, which may contain a more diverse population of H. influenzae than nasopharyngeal cultures (8). Alternatively, these findings could be associated with our the bacterial culture and genetic typing techniques. We used selective media for the initial bacterial isolation, chose 10 colonies from the plates for further examination, and used PFGE to type all isolates. Use of PFGE, the most sensitive method available for genetically typing bacteria, may have resulted in detection of more genetic differences than other commonly used typing methods, such as multilocus enzyme electrophoresis (8).
To address the possibility that colonization with multiple genetic types was due to colonization with a single isolate that was genetically evolving during the study period, we defined strain identity according to the criteria of Tenover et al. (14). Few nonidentical PFGE patterns in our study appeared related; most had seven or more restriction fragment differences, which implies at least three genetic differences. Although the evolutionary dynamics that have resulted in a fairly high diversity among several surface exposed proteins of H. influenzae (21) have not been well defined, the large differences we observed in the PFGE patterns are unlikely to have occurred by evolutionary changes over the 1-week sampling intervals.
The association between risk factors for otitis media and colonization with multiple genetic types of bacteria had not been examined before this study. We found that children who were exposed to smoking were more likely to carry multiple H. influenzae isolates than children who were not. Smoking has been described as a risk factor for otitis media (9,11,12,22,23), but the exact mechanism by which it contributes to otitis media had not been described. Mucociliary clearance is damaged in adult smokers (24), and H. influenzae is more likely to adhere to pharyngeal cells of middle-aged smokers than nonsmokers with bronchitis (25). Etzel et al. (22) suggested that secondhand smoke may also damage the upper respiratory environment in young children, leading to increased bacterial colonization. Increased carriage of diverse H. influenzae types may partially account for the increased episodes of otitis media observed in children exposed to smoking. However, the association of smoking with both increased carriage of H. influenzae and episodes of otitis media does not establish a causal relationship.
Children with allergies were more likely to carry multiple H. influenzae isolates than children without allergies. Allergies in young children have also been described as a risk factor for otitis media (12,26-28), because upper respiratory mucosal swelling during an allergic episode may cause eustachian tube dysfunction, similar to that observed during an upper respiratory viral infection (26,28). Alternatively, allergic responses may result in impaired mucociliary activity (29,30), which may permit increased bacterial colonization of the upper respiratory tract. As with smoking, while allergies are associated with both increased carriage of H. influenzae and increased episodes of otitis media, increased carriage of H. influenzae does not necessarily increase episodes of otitis media.
Children 36 to 47 months of age were more likely to carry multiple types of H. influenzae than children >23 months of age. The reason for this association is unclear. Children in the 36- to 47-month age group were not significantly more likely to have specific otitis media risk factors than younger children. However, 36- to 47-month-old children may have more contacts than younger children, which could contribute to increased exposure to multiple genetic types of H. influenzae.
Children who were given an antibiotic during the study period were less likely to carry multiple types of H. influenzae than children who were not. As antibiotic use reduces or eliminates bacterial carriage, this factor may explain why children who took an antibiotic were less likely to carry multiple H. influenzae isolates.
No associations were found between risk factors for otitis media and colonization with multiple genetic types of S. pneumoniae. Only six of the throat culture specimens contained multiple S. pneumoniae isolates, and such a small sample size would probably not be sufficient to detect any associations; five of the six cultures also contained multiple genetic types of H. influenzae. Therefore, some children appear more likely to carry multiple types of both bacterial species.
Many parents were reluctant to allow their children to participate in our study (49% participation overall); thus, our study population may not accurately reflect the entire day-care population. Parents whose children had a history of frequent otitis media may have been more willing to participate. These children might also have been more likely to be exposed to smoking or have allergies and may have also carried more types of H. influenzae. However, we believe this bias is unlikely, as we found no association between carriage of multiple H. influenzae and history of frequent ear infections. Alternatively, parents who had other otitis media-prone children may have been more concerned about otitis media in general and more likely to allow their other children to participate. The participating children might therefore have been exposed to the same risk factors as their otitis-prone siblings and have been more likely to carry multiple H. influenzae isolates. This is also unlikely, as children with siblings were no more likely to carry multiple isolates than children without siblings. Therefore, the associations between specific risk factors for otitis media and colonization with multiple bacterial strains observed may be a real phenomenon in day-care populations.
We may also have been limited in our ability to detect colonizing S. pneumoniae. Pharyngeal swabs are less effective for isolation of S. pneumoniae than nasopharyngeal swabs (31,32); we may have missed approximately 13% of colonizing S. pneumoniae isolates. In addition, all isolates that were typed by optochin disc sensitivity had zones of inhibition at least 17 mm. (A zone of inhibition at least 14 mm is necessary for presumptive identification of S. pneumoniae.) We did not type (by PFGE) any potential isolates with a smaller zone of inhibition; therefore, we may have missed some isolates that would have yielded a questionable inhibition zone. These limitations in our sampling and culture techniques could have resulted in an underestimate of the genetic diversity of the S. pneumoniae isolates carried by this group of children.
In summary, this study describes high genetic diversity among S. pneumoniae and H. influenzae isolates colonizing individual children attending two day-care centers. Carriage of multiple genetic types of H. influenzae was associated with child's age, antibiotic use, exposure to smoking, and history of allergies. Carriage of multiple genetic types of H. influenzae may be an intermediate step in the biologic pathway leading to otitis media and may explain the increased episodes of otitis media observed in children with allergies or children exposed to smoking. Future studies will determine 1) if the high rate of carriage of multiple H. influenzae is specific to children attending day-care centers or if all children commonly carry multiple H. influenzae pathogens in their upper respiratory tracts and 2) if children carrying multiple pathogens are also at increased risk for otitis media.
Dr. St. Sauver is with the Mayo Clinic and Foundation, Department of Health Sciences Research, Section of Clinical Epidemiology, Rochester, Minnesota. Her research interests include the epidemiology of pediatric infectious diseases.
Acknowledgment
We thank Valerie Reed for help with the bacterial isolation and identification protocols, Patricia Tallman for assistance in isolating the bacteria, and Chi Ching Chang for the PFGE protocol; and Susan Murray, Rosemary Rochford, and Mark Wilson for their review of the manuscript.
References
- Gates GA. Cost-effectiveness considerations in otitis media treatment. Otolaryngol Head Neck Surg. 1996;114:525–30. DOIPubMedGoogle Scholar
- Henderson FW, Collier AM, Sanyal MA, Watkins JM, Fairclough DL, Clyde WA Jr, A longitudinal study of respiratory viruses and bacteria in the etiology of acute otitis media with effusion. N Engl J Med. 1982;306:1377–83. DOIPubMedGoogle Scholar
- Harabuchi Y, Faden H, Yamanaka N, Duffy L, Wolf J, Krystofik D. Human milk secretory IgA antibody to nontypeable Haemophilus influenzae: possible protective effects against nasopharyngeal colonization. J Pediatr. 1994;124:193–8. DOIPubMedGoogle Scholar
- Leach AJ, Boswell JB, Asche V, Nienhuys TG, Mathews JD. Bacterial colonization of the nasopharynx predicts very early onset and persistence of otitis media in Australian aboriginal infants. Pediatr Infect Dis J. 1994;13:983–9. DOIPubMedGoogle Scholar
- Harabuchi Y, Faden H, Yamanaka N, Duffy L, Wolf J, Krystofik D, Nasopharyngeal colonization with nontypeable Haemophilus influenzae and recurrent otitis media. J Infect Dis. 1994;170:862–6.PubMedGoogle Scholar
- Faden H, Duffy L, Wasielewski R, Wolf J, Krystofik YT. Tonawanda/Williamsville Pediatrics. Relationship between nasopharyngeal colonization and the development of otitis media in children. J Infect Dis. 1997;175:1440–5. DOIPubMedGoogle Scholar
- Smith-Vaughan HC, Leach AJ, Shelby-James TM, Kemp K, Kemp DJ, Mathews JD. Carriage of multiple ribotypes of nonencapsulated Haemophilus influenzae in aboriginal infants with otitis media. Epidemiol Infect. 1996;116:177–83. DOIPubMedGoogle Scholar
- Alho OP, Laara E, Oja H. Public health impact of various risk factors for acute otitis media in northern Finland. Am J Epidemiol. 1996;143:1149–56.PubMedGoogle Scholar
- Uhari M, Mantysaari K, Niemela M. A meta-analytic review of the risk factors for acute otitis media. Clin Infect Dis. 1996;22:1079–83.PubMedGoogle Scholar
- Paradise JL, Rockette HE, Colborn DK, Bernard BS, Smith CG, Kurs-Lasky M, Otitis media in 2253 Pittsburgh-area infants: prevalence and risk factors during the first two years of life. Pediatrics. 1997;99:318–33. DOIPubMedGoogle Scholar
- Stenstrom C, Ingvarsson L. Otitis-prone children and controls: a study of possible predisposing factors. 2. Physical findings, frequency of illness, allergy, day care, and parental smoking. Acta Otolaryngol. 1997;117:696–703. DOIPubMedGoogle Scholar
- Gray BM, Converse GM III, Dillon HC Jr. Epidemiologic studies of Streptococcus pneumoniae in infants: acquisition, carriage, and infection during the first 24 months of life. J Infect Dis. 1980;142:923–33.PubMedGoogle Scholar
- Tenover FC, Arbeit RD, Goering RV, Mickelsen AP, Murray BE, Persing DH, Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233–9.PubMedGoogle Scholar
- Adobe Systems Inc. Adobe Photoshop. Version 4.0, San Jose (CA): Adobe Systems;1997.
- SAS Institute, Inc. SAS. Version 6.12, Cary (NC): SAS Institute; 1995.
- Moller LV, Regelink AG, Grasselier H, Dankert-Roelse JE, Dankert J, van Alphen L. Multiple Haemophilus influenzae strains and strain variants coexist in the respiratory tracts of patients with cystic fibrosis. J Infect Dis. 1995;172:1388–92.PubMedGoogle Scholar
- Murphy TF, Sethi S, Klingman KL, Bruggemann AB, Doern GV. Simultaneous respiratory tract colonization by multiple strains of nontypeable Haemophilus influenzae in chronic obstructive pulmonary disease: implications for antibiotic therapy. J Infect Dis. 1999;180:404–9. DOIPubMedGoogle Scholar
- Murphy TF, Bernstein JM, Dryja DM, Campagnari AA, Apicella MA. Outer membrane protein and lipooligosaccharide analysis of paired nasopharyngeal and middle ear isolates in otitis media due to nontypeable Haemophilus influenzae: pathogenic and epidemiological observations. J Infect Dis. 1987;156:723–31.PubMedGoogle Scholar
- Bernstein JM, Dryja D, Yuskiw N, Murphy TF. Analysis of isolates recovered from multiple sites of the nasopharynx of children colonized by nontypeable Haemophilus influenzae. Eur J Clin Microbiol Infect Dis. 1997;16:750–3. DOIPubMedGoogle Scholar
- Gilsdorf JR. Antigenic diversity and gene polymorphisms in Haemophilus influenzae. Infect Immun. 1998;66:5053–9.PubMedGoogle Scholar
- Etzel RA, Pattishall EN, Haley NJ, Fletcher RH, Henderson FW. Passive smoking and middle ear effusion among children in day care. Pediatrics. 1992;90:228–32.PubMedGoogle Scholar
- Teele DW, Klein JO, Rosner B. Epidemiology of otitis media during the first seven years of life in children in greater Boston: a prospective, cohort study. J Infect Dis. 1989;160:83–94.PubMedGoogle Scholar
- Vastag E, Matthys H, Zsamboki G, Kohler D, Daikeler G. Mucociliary clearance in smokers. Eur J Respir Dis. 1986;68:107–13.PubMedGoogle Scholar
- Fainstein V, Musher DM. Bacterial adherence to pharyngeal cells in smokers, nonsmokers, and chronic bronchitics. Infect Immun. 1979;26:178–82.PubMedGoogle Scholar
- Fireman P. Otitis media and eustachian tube dysfunction: connection to allergic rhinitis. J Allergy Clin Immunol. 1997;99:S787–97. DOIPubMedGoogle Scholar
- Marx J, Osguthorpe JD, Parsons G. Day care and the incidence of otitis media in young children. Otolaryngol Head Neck Surg. 1995;112:695–9. DOIPubMedGoogle Scholar
- Kvaerner KJ, Tambs K, Harris JR, Mair IW, Magnus P. Otitis media: relationship to tonsillitis, sinusitis, and atopic diseases. Int J Pediatr Otorhinolaryngol. 1996;35:127–41. DOIPubMedGoogle Scholar
- Hastie AT, Everts KB, Zangrilli J, Shaver JR, Pollice MB, Fish JE, HSP27 elevated in mild allergic inflammation protects airway epithelium from H2SO4 effects. Am J Physiol. 1997;273:L401–9.PubMedGoogle Scholar
- Tanaka A, Ohashi Y, Kakinoki Y, Washio Y, Kishimoto K, Ohno Y, Influence of the allergic response on the mucociliary system in the eustachian tube. Acta Otolaryngol Suppl (Stockh). 1998;538:98–101. DOIPubMedGoogle Scholar
- Gray BM, Converse GM III, Dillon HC Jr. Epidemiologic studies of Streptococcus pneumoniae in infants: acquisition, carriage, and infection during the first 24 months of life. J Infect Dis. 1980;142:923–33.PubMedGoogle Scholar
- Rapola S, Salo E, Kiiski P. Leinonen, Takala AK. Comparison of four different sampling methods for detecting pharyngeal carriage of Streptococcus pneumoniae and Haemophilus influenzae in children. J Clin Microbiol. 1997;35:1077–9.PubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 6, Number 6—December 2000
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Janet Gilsdorf, Department of Pediatrics and Communicable Diseases, L2225 Women's Hospital, Box 0244, University of Michigan Medical Center, Ann Arbor, MI, 48109-0244; Fax: 734-936-7635
Top